

The N-terminal domain of mammalian soluble epoxide hydrolase is a phosphatase
- PMID: 12574508
- PMCID: PMC149870
- DOI: 10.1073/pnas.0437829100
The N-terminal domain of mammalian soluble epoxide hydrolase is a phosphatase
Abstract
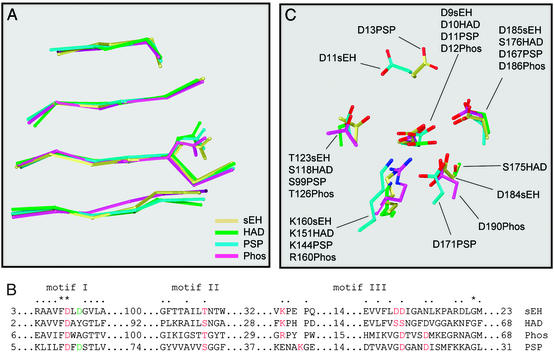
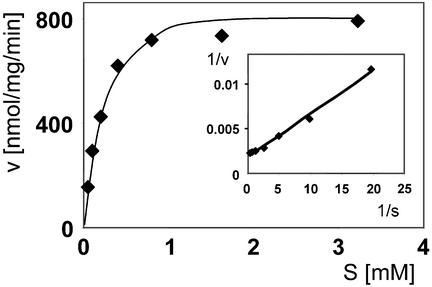
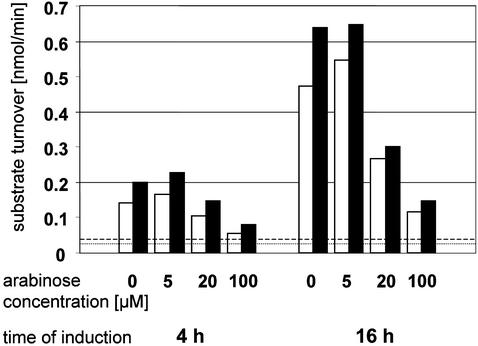
The mammalian soluble epoxide hydrolase (sEH) is an enzyme with multiple functions, being implicated in detoxification of xenobiotic epoxides as well as in regulation of physiological processes such as blood pressure. The enzyme is a homodimer, in which each subunit is composed of two domains. The 35-kDa C-terminal domain has an alpha/beta hydrolase fold and harbors the catalytic center for the EH activity. The 25-kDa N-terminal domain has a different alpha/beta fold and belongs to the haloacid dehalogenase superfamily of enzymes. The catalytic properties of the enzyme reported so far can all be explained by the action of the C-terminal domain alone. The function of the N-terminal domain, other than in structural stabilization of the dimer, has therefore remained unclear. By structural comparison of this domain to other haloacid dehalogenase family members, we identified a putative active site containing all necessary components for phosphatase activity. Subsequently, we found rat sEH hydrolyzed 4-nitrophenyl phosphate with a rate constant of 0.8 s(-1) and a K(m) of 0.24 mM. Recombinant human sEH lacking the C-terminal domain also displayed phosphatase activity. Presence of a phosphatase substrate did not affect epoxide turnover nor did epoxides affect dephosphorylation by the intact enzyme, indicating both catalytic sites act independently. The enzyme was unable to hydrolyze 4-nitrophenyl sulfate, suggesting its role in xenobiotic metabolism does not extend beyond phosphates. Thus, we propose this domain participates instead in the regulation of the physiological functions associated with sEH.
Figures
References
-
- Arand M, Oesch F. In: Enzyme Systems That Metabolise Drugs and Other Xenobiotics. Ioannides C, editor. New York: Wiley; 2002. pp. 459–483.
-
- Oesch F. Xenobiotica. 1973;3:305–340. - PubMed
-
- Yu Z, Xu F, Huse L M, Morisseau C, Draper A J, Newman J W, Parker C, Graham L, Engler M M, Hammock B D, et al. Circ Res. 2000;87:992–998. - PubMed
-
- Sinal C J, Miyata M, Tohkin M, Nagata K, Bend J R, Gonzalez F J. J Biol Chem. 2000;275:40504–40510. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
