

Distinct contributions of small and large conductance Ca2+-activated K+ channels to rat Purkinje neuron function
- PMID: 12576503
- PMCID: PMC2342800
- DOI: 10.1113/jphysiol.2002.027854
Distinct contributions of small and large conductance Ca2+-activated K+ channels to rat Purkinje neuron function
Abstract
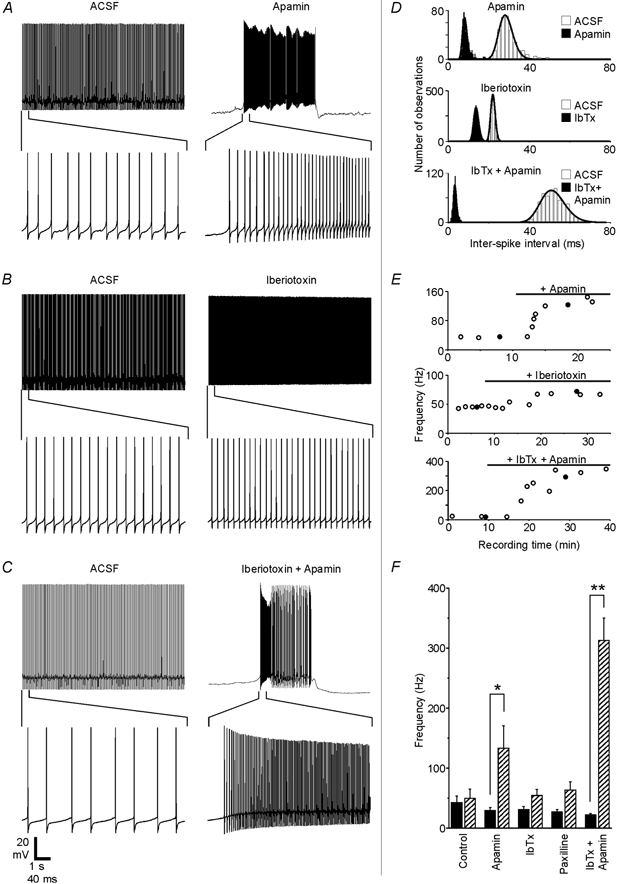
The cerebellum is important for many aspects of behaviour, from posture maintenance and goal-oriented reaching movements to timing tasks and certain forms of learning. In every case, information flowing through the cerebellum passes through Purkinje neurons, which receive input from the two primary cerebellar afferents and generate continuous streams of action potentials that constitute the sole output from the cerebellar cortex to the deep nuclei. The tonic firing behaviour observed in Purkinje neurons in vivo is maintained in brain slices even when synaptic inputs are blocked, suggesting that Purkinje neuron activity relies to a significant extent on intrinsic conductances. Previous research has suggested that the interplay between Ca2+ currents and Ca2+-activated K+ channels (KCa channels) is important for Purkinje cell activity, but how many different KCa channel types are present and what each channel type contributes to cell behaviour remains unclear. In order to better understand the ionic mechanisms that control the behaviour of these neurons, we investigated the effects of different Ca2+ channel and KCa channel antagonists on Purkinje neurons in acute slices of rat cerebellum. Our data show that Ca2+ entering through P-type voltage-gated Ca2+ channels activates both small-conductance (SK) and large-conductance (BK) KCa channels. SK channels play a role in setting the intrinsic firing frequency, while BK channels regulate action potential shape and may contribute to the unique climbing fibre response.
Figures






References
-
- Blatz AL, Magleby KL. Single apamin-blocked Ca-activated K+ channels of small conductance in cultured rat skeletal muscle. Nature. 1986;323:718–720. - PubMed
-
- Chung YH, Shin C, Park KH, Cha CI. Immunohistochemical study on the distribution of neuronal voltage-gated calcium channels in the rat cerebellum. Brain Res. 2000;865:278–282. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous