

Cyclic AMP induces integrin-mediated cell adhesion through Epac and Rap1 upon stimulation of the beta 2-adrenergic receptor
- PMID: 12578910
- PMCID: PMC2173739
- DOI: 10.1083/jcb.200209105
Cyclic AMP induces integrin-mediated cell adhesion through Epac and Rap1 upon stimulation of the beta 2-adrenergic receptor
Abstract
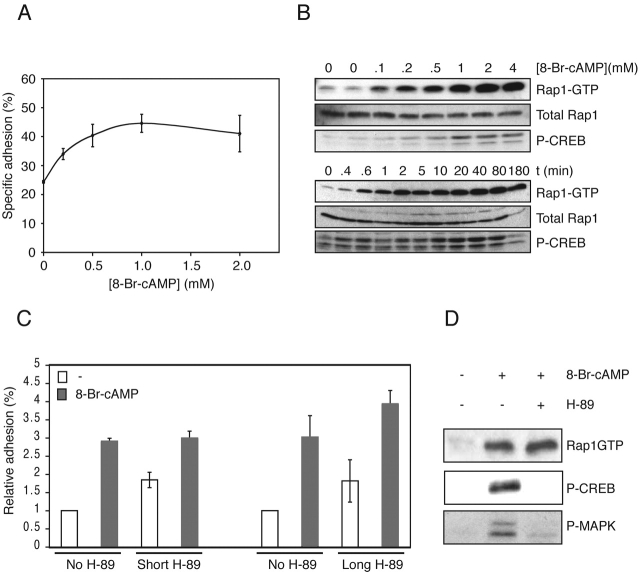
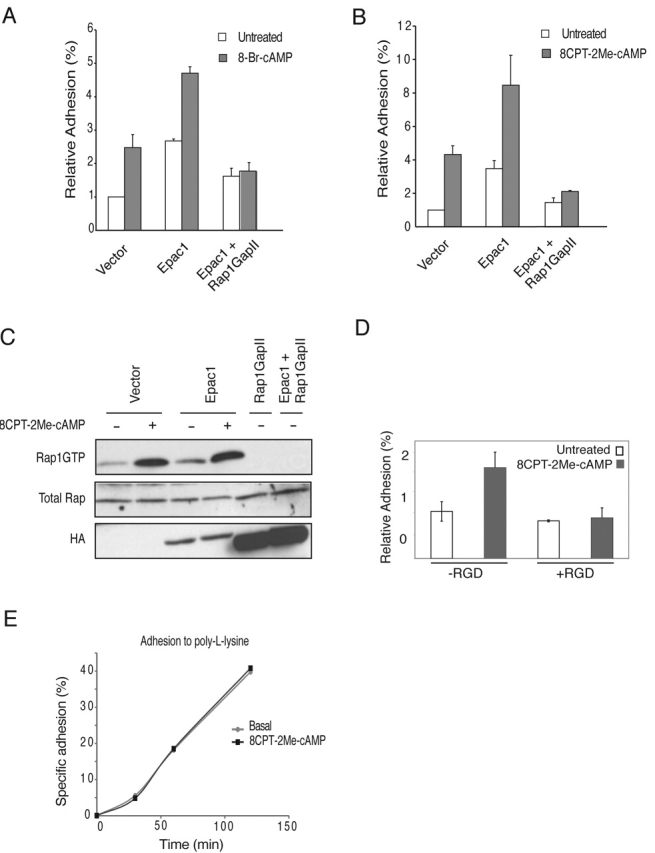
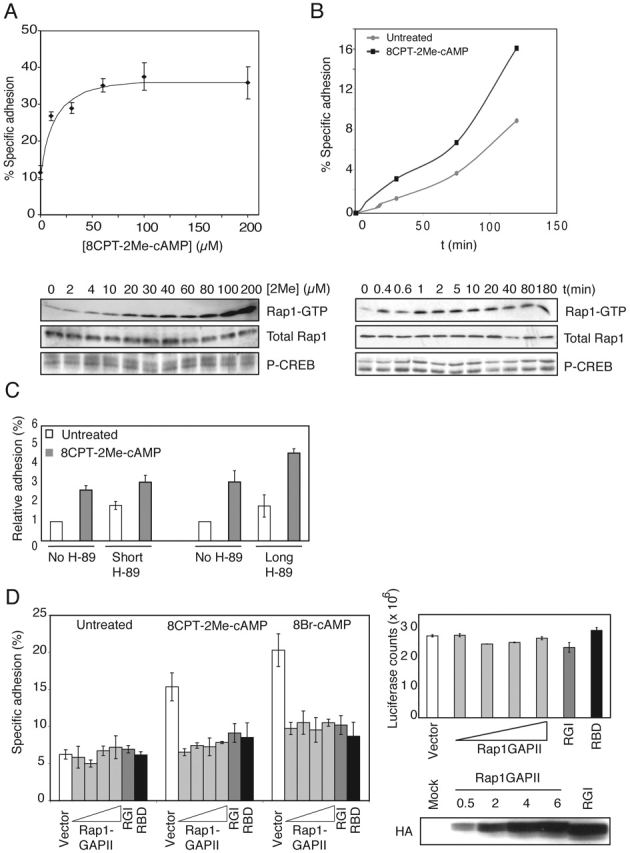
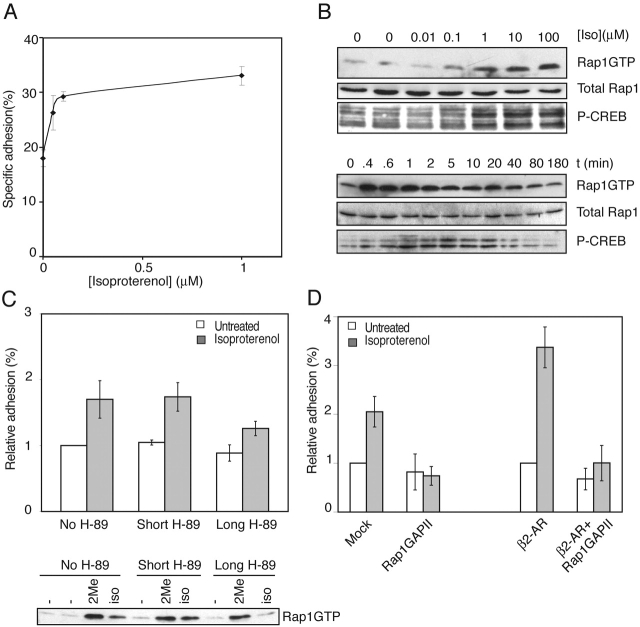
cAMP controls many cellular processes mainly through the activation of protein kinase A (PKA). However, more recently PKA-independent pathways have been established through the exchange protein directly activated by cAMP (Epac), a guanine nucleotide exchange factor for the small GTPases Rap1 and Rap2. In this report, we show that cAMP can induce integrin-mediated cell adhesion through Epac and Rap1. Indeed, when Ovcar3 cells were treated with cAMP, cells adhered more rapidly to fibronectin. This cAMP effect was insensitive to the PKA inhibitor H-89. A similar increase was observed when the cells were transfected with Epac. Both the cAMP effect and the Epac effect on cell adhesion were abolished by the expression of Rap1-GTPase-activating protein, indicating the involvement of Rap1 in the signaling pathway. Importantly, a recently characterized cAMP analogue, 8-(4-chloro-phenylthio)-2'-O-methyladenosine-3',5'-cyclic monophosphate, which specifically activates Epac but not PKA, induced Rap-dependent cell adhesion. Finally, we demonstrate that external stimuli of cAMP signaling, i.e., isoproterenol, which activates the G alpha s-coupled beta 2-adrenergic receptor can induce integrin-mediated cell adhesion through the Epac-Rap1 pathway. From these results we conclude that cAMP mediates receptor-induced integrin-mediated cell adhesion to fibronectin through the Epac-Rap1 signaling pathway.
Figures
References
-
- Arai, A., Y. Nosaka, E. Kanda, K. Yamamoto, N. Miyasaka, and O. Miura. 2001. Rap1 is activated by erythropoietin or interleukin-3 and is involved in regulation of β1 integrin-mediated hematopoietic cell adhesion. J. Biol. Chem. 276:10453–10462. - PubMed
-
- Bertoni, A., S. Tadokoro, K. Eto, N. Pampori, L.V. Parise, G.C. White, and S.J. Shattil. 2002. Relationships between Rap1b, affinity modulation of integrin α IIbβ 3, and the actin cytoskeleton. J. Biol. Chem. 277:25715–25721. - PubMed
-
- Bos, J.L., J. de Rooij, and K.A. Reedquist. 2001. Rap1 signalling: adhering to new models. Nat. Rev. Mol. Cell Biol. 2:369–377. - PubMed
-
- Buczek-Thomas, J.A., N. Chen, and T. Hasan. 1998. Integrin-mediated adhesion and signalling in ovarian cancer cells. Cell. Signal. 10:55–63. - PubMed
