

Quantitative role of the human papillomavirus type 16 E5 gene during the productive stage of the viral life cycle
- PMID: 12584306
- PMCID: PMC149772
- DOI: 10.1128/jvi.77.5.2832-2842.2003
Quantitative role of the human papillomavirus type 16 E5 gene during the productive stage of the viral life cycle
Abstract
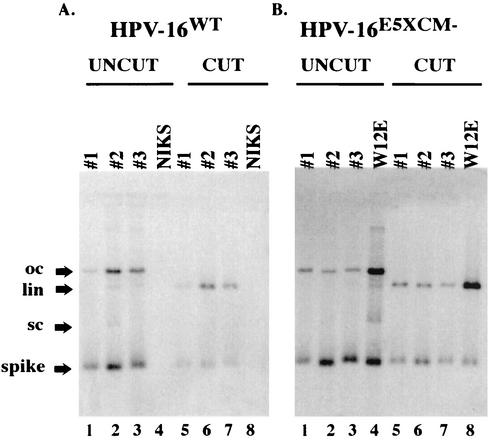
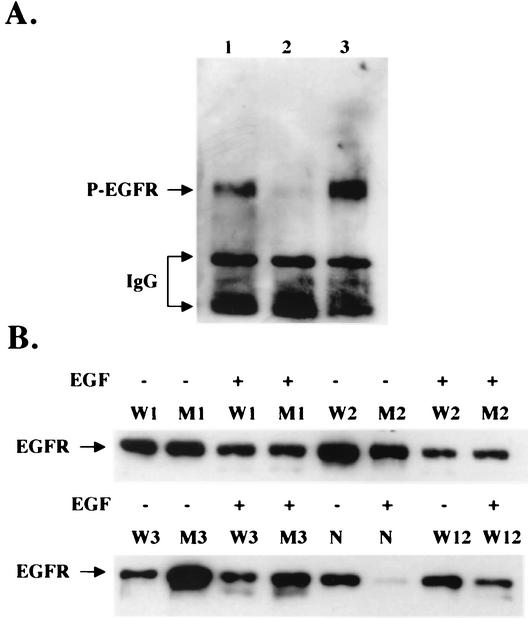
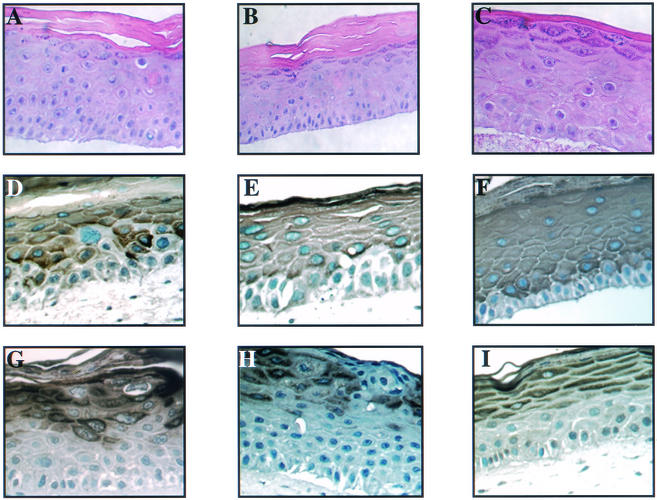
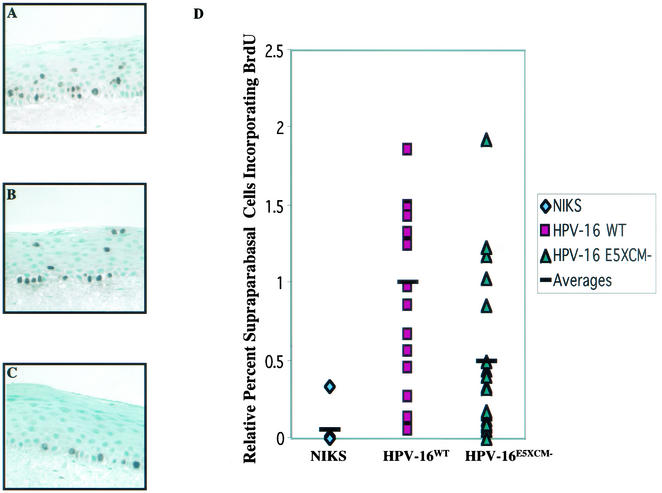
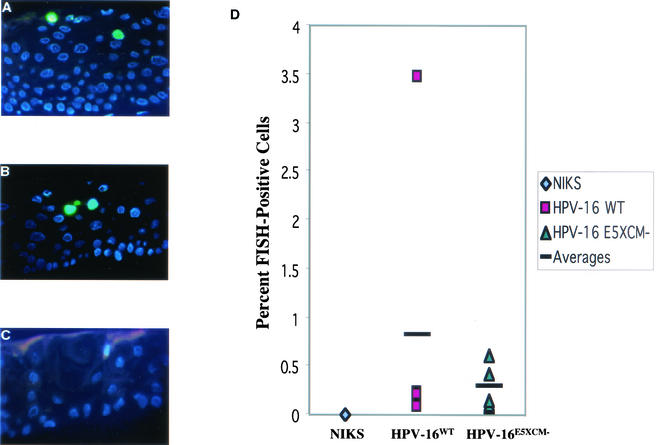
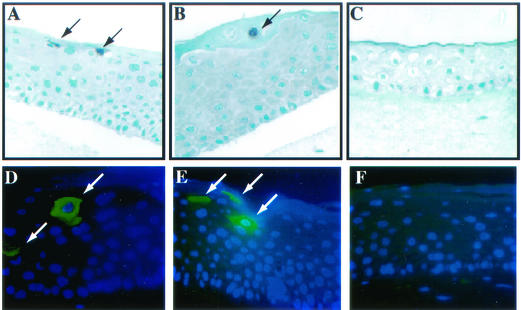
Human papillomaviruses (HPVs) are small circular DNA viruses that cause warts. Infection with high-risk anogenital HPVs, such as HPV type 16 (HPV16), is associated with human cancers, specifically cervical cancer. The life cycle of HPVs is intimately tied to the differentiation status of the host epithelium and has two distinct stages: the nonproductive stage and the productive stage. In the nonproductive stage, which arises in the poorly differentiated basal epithelial compartment of a wart, the virus maintains itself as a low-copy-number nuclear plasmid. In the productive stage, which arises as the host cell undergoes terminal differentiation, viral DNA is amplified; the capsid genes, L1 and L2, are expressed; and progeny virions are produced. This stage of the viral life cycle relies on the ability of the virus to reprogram the differentiated cells to support DNA synthesis. Papillomaviruses encode multiple oncoproteins, E5, E6, and E7. In the present study, we analyze the role of one of these viral oncogenes, E5, in the viral life cycle. To assess the role of E5 in the HPV16 life cycle, we introduced wild-type (WT) or E5 mutant HPV16 genomes into NIKS, a keratinocyte cell line that supports the papillomavirus life cycle. By culturing these cells under conditions that allow them to remain undifferentiated, a state similar to that of basal epithelial cells, we determined that E5 does not play an essential role in the nonproductive stage of the HPV16 life cycle. To determine if E5 plays a role in the productive stage of the viral life cycle, we cultured keratinocyte populations in organotypic raft cultures, which promote the differentiation and stratification of epithelial cells. We found that cells harboring E5 mutant genomes displayed a quantitative reduction in the percentage of suprabasal cells undergoing DNA synthesis, compared to cells containing WT HPV16 DNA. This reduction in DNA synthesis, however, did not prevent amplification of viral DNA in the differentiated cellular compartment. Likewise, late viral gene expression and the perturbation of normal keratinocyte differentiation were retained in cells harboring E5 mutant genomes. These data demonstrate that E5 plays a subtle role during the productive stage of the HPV16 life cycle.
Figures


Similar articles
-
The human papillomavirus type 16 E7 oncogene is required for the productive stage of the viral life cycle.J Virol. 2000 Jul;74(14):6622-31. doi: 10.1128/jvi.74.14.6622-6631.2000. J Virol. 2000. PMID: 10864676 Free PMC article.
-
Human papillomavirus type 31 E5 protein supports cell cycle progression and activates late viral functions upon epithelial differentiation.J Virol. 2003 Mar;77(5):2819-31. doi: 10.1128/jvi.77.5.2819-2831.2003. J Virol. 2003. PMID: 12584305 Free PMC article.
-
Establishment of the human papillomavirus type 16 (HPV-16) life cycle in an immortalized human foreskin keratinocyte cell line.Virology. 1999 Sep 30;262(2):344-54. doi: 10.1006/viro.1999.9868. Virology. 1999. PMID: 10502513
-
Pathogenesis of human papillomaviruses in differentiating epithelia.Microbiol Mol Biol Rev. 2004 Jun;68(2):362-72. doi: 10.1128/MMBR.68.2.362-372.2004. Microbiol Mol Biol Rev. 2004. PMID: 15187189 Free PMC article. Review.
-
[Virological and carcinogenic aspects of HPV].Bull Acad Natl Med. 2007 Mar;191(3):611-23; discussion 623. Bull Acad Natl Med. 2007. PMID: 18072657 Review. French.
Cited by
-
The minor capsid protein L2 contributes to two steps in the human papillomavirus type 31 life cycle.J Virol. 2005 Apr;79(7):3938-48. doi: 10.1128/JVI.79.7.3938-3948.2005. J Virol. 2005. PMID: 15767396 Free PMC article.
-
Human papillomavirus 16 E5 modulates the expression of host microRNAs.PLoS One. 2011;6(7):e21646. doi: 10.1371/journal.pone.0021646. Epub 2011 Jul 1. PLoS One. 2011. PMID: 21747943 Free PMC article.
-
Tumourigenesis driven by the human papillomavirus type 16 Asian-American e6 variant in a three-dimensional keratinocyte model.PLoS One. 2014 Jul 1;9(7):e101540. doi: 10.1371/journal.pone.0101540. eCollection 2014. PLoS One. 2014. PMID: 24983759 Free PMC article.
-
Genomic Instability and DNA Damage Repair Pathways Induced by Human Papillomaviruses.Viruses. 2021 Sep 14;13(9):1821. doi: 10.3390/v13091821. Viruses. 2021. PMID: 34578402 Free PMC article. Review.
-
Infectious titres of human papillomaviruses (HPVs) in patient lesions, methodological considerations in evaluating HPV infectivity and implications for the efficacy of high-level disinfectants.EBioMedicine. 2021 Jan;63:103165. doi: 10.1016/j.ebiom.2020.103165. Epub 2021 Jan 7. EBioMedicine. 2021. PMID: 33422988 Free PMC article.
References
-
- Adam, J. L., M. W. Briggs, and D. J. McCance. 2000. A mutagenic analysis of the E5 protein of human papillomavirus type 16 reveals that E5 binding to the vacuolar H+-ATPase is not sufficient for biological activity, using mammalian and yeast expression systems. Virology 272:315-325. - PubMed
-
- Allen-Hoffman, B. L., S. L. Schlosser, C. A. Ivarie, L. F. Meisner, and O. C. S. L. 2000. Normal growth and differentiation in a spontaneously immortalized near-diploid human keratinocyte cell line, NIKS. J. Investig. Dermatol. 114:444-455. - PubMed
-
- Banks, L., C. Edmonds, and K. H. Vousden. 1990. Ability of the HPV16 E7 protein to bind RB and induce DNA synthesis is not sufficient for efficient transforming activity in NIH3T3 cells. Oncogene 5:1383-1389. - PubMed
-
- Bouvard, V., G. Matlashewski, Z. M. Gu, A. Storey, and L. Banks. 1994. The human papillomavirus type 16 E5 gene cooperates with the E7 gene to stimulate proliferation of primary cells and increases viral gene expression. Virology 203:73-80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
