

Mutation of host DnaJ homolog inhibits brome mosaic virus negative-strand RNA synthesis
- PMID: 12584324
- PMCID: PMC149758
- DOI: 10.1128/jvi.77.5.2990-2997.2003
Mutation of host DnaJ homolog inhibits brome mosaic virus negative-strand RNA synthesis
Abstract
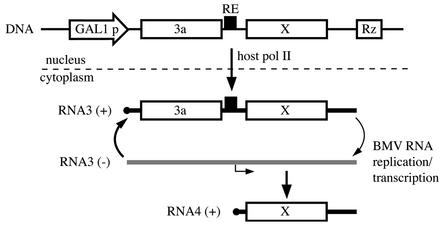
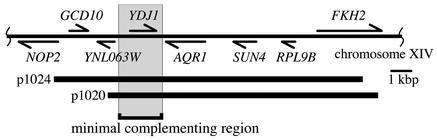
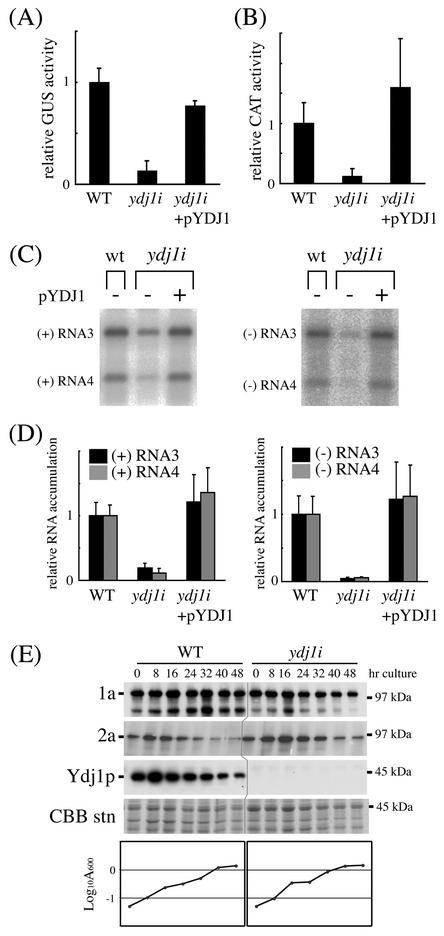
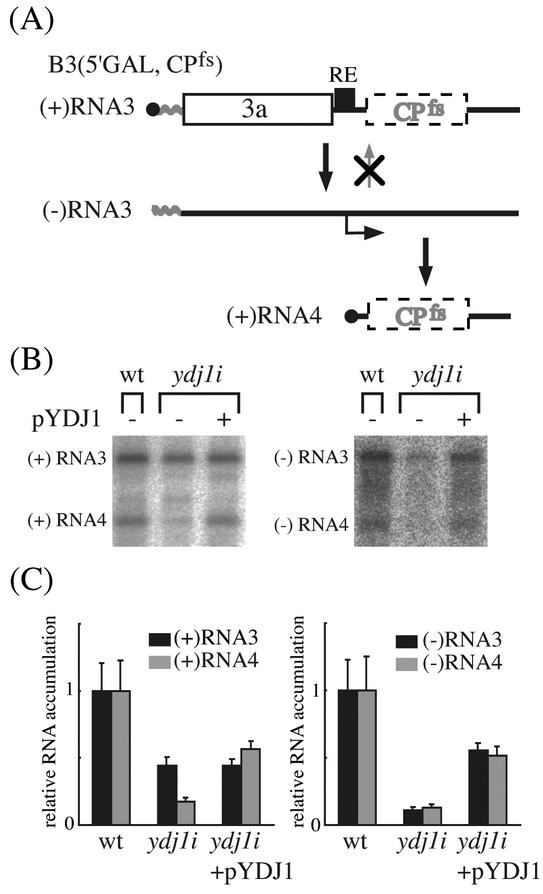
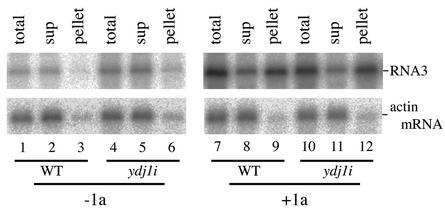
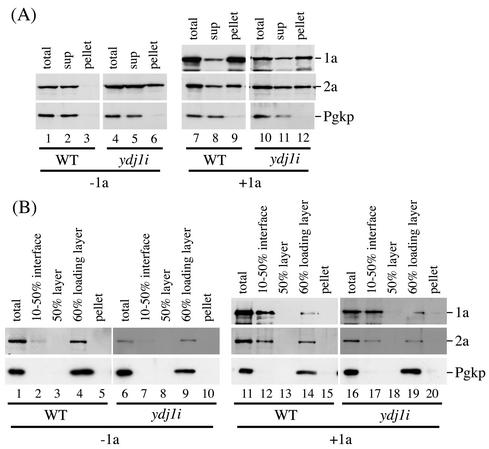
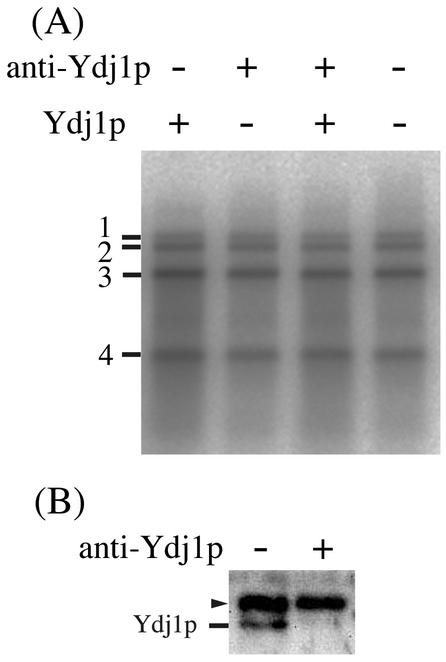
The replication of positive-strand RNA viruses involves not only viral proteins but also multiple cellular proteins and intracellular membranes. In both plant cells and the yeast Saccharomyces cerevisiae, brome mosaic virus (BMV), a member of the alphavirus-like superfamily, replicates its RNA in endoplasmic reticulum (ER)-associated complexes containing viral 1a and 2a proteins. Prior to negative-strand RNA synthesis, 1a localizes to ER membranes and recruits both positive-strand BMV RNA templates and the polymerase-like 2a protein to ER membranes. Here, we show that BMV RNA replication in S. cerevisiae is markedly inhibited by a mutation in the host YDJ1 gene, which encodes a chaperone Ydj1p related to Escherichia coli DnaJ. In the ydj1 mutant, negative-strand RNA accumulation was inhibited even though 1a protein associated with membranes and the positive-strand RNA3 replication template and 2a protein were recruited to membranes as in wild-type cells. In addition, we found that in ydj1 mutant cells but not wild-type cells, a fraction of 2a protein accumulated in a membrane-free but insoluble, rapidly sedimenting form. These and other results show that Ydj1p is involved in forming BMV replication complexes active in negative-strand RNA synthesis and suggest that a chaperone system involving Ydj1p participates in 2a protein folding or assembly into the active replication complex.
Figures
Similar articles
-
Brome mosaic virus RNA replication proteins 1a and 2a colocalize and 1a independently localizes on the yeast endoplasmic reticulum.J Virol. 1999 Dec;73(12):10303-9. doi: 10.1128/JVI.73.12.10303-10309.1999. J Virol. 1999. PMID: 10559348 Free PMC article.
-
Yeast mutations in multiple complementation groups inhibit brome mosaic virus RNA replication and transcription and perturb regulated expression of the viral polymerase-like gene.Proc Natl Acad Sci U S A. 1997 Dec 9;94(25):13810-5. doi: 10.1073/pnas.94.25.13810. Proc Natl Acad Sci U S A. 1997. PMID: 9391109 Free PMC article.
-
An alternate pathway for recruiting template RNA to the brome mosaic virus RNA replication complex.J Virol. 2003 Feb;77(4):2568-77. doi: 10.1128/jvi.77.4.2568-2577.2003. J Virol. 2003. PMID: 12551995 Free PMC article.
-
Bromovirus-induced remodeling of host membranes during viral RNA replication.Curr Opin Virol. 2014 Dec;9:104-10. doi: 10.1016/j.coviro.2014.09.018. Epub 2014 Oct 16. Curr Opin Virol. 2014. PMID: 25462441 Review.
-
Brome mosaic virus RNA replication: revealing the role of the host in RNA virus replication.Annu Rev Phytopathol. 2003;41:77-98. doi: 10.1146/annurev.phyto.41.052002.095717. Epub 2003 Mar 10. Annu Rev Phytopathol. 2003. PMID: 12651962 Review.
Cited by
-
Viral and host determinants of RNA virus vector replication and expression.Vaccine. 2005 Mar 7;23(15):1784-7. doi: 10.1016/j.vaccine.2004.11.005. Vaccine. 2005. PMID: 15734041 Free PMC article. Review.
-
Influence of cytoplasmic heat shock protein 70 on viral infection of Nicotiana benthamiana.Mol Plant Pathol. 2008 Nov;9(6):809-17. doi: 10.1111/j.1364-3703.2008.00505.x. Mol Plant Pathol. 2008. PMID: 19019009 Free PMC article.
-
A targeted analysis of cellular chaperones reveals contrasting roles for heat shock protein 70 in flock house virus RNA replication.J Virol. 2010 Jan;84(1):330-9. doi: 10.1128/JVI.01808-09. J Virol. 2010. PMID: 19828623 Free PMC article.
-
Alternate, virus-induced membrane rearrangements support positive-strand RNA virus genome replication.Proc Natl Acad Sci U S A. 2004 Aug 3;101(31):11263-8. doi: 10.1073/pnas.0404157101. Epub 2004 Jul 27. Proc Natl Acad Sci U S A. 2004. PMID: 15280537 Free PMC article.
-
A vicilin-like seed storage protein, PAP85, is involved in tobacco mosaic virus replication.J Virol. 2013 Jun;87(12):6888-900. doi: 10.1128/JVI.00268-13. Epub 2013 Apr 10. J Virol. 2013. PMID: 23576511 Free PMC article.
References
-
- Ahlquist, P. 1992. Bromovirus RNA replication and transcription. Curr. Opin. Genet. Dev. 2:71-76. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl (ed.). 1989. Current protocols in molecular biology. John Wiley and Sons, New York, N.Y.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
