

Effects of point mutations in the major capsid protein of beet western yellows virus on capsid formation, virus accumulation, and aphid transmission
- PMID: 12584348
- PMCID: PMC149785
- DOI: 10.1128/jvi.77.5.3247-3256.2003
Effects of point mutations in the major capsid protein of beet western yellows virus on capsid formation, virus accumulation, and aphid transmission
Abstract
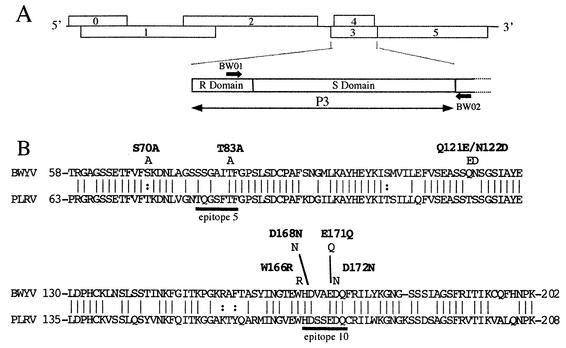
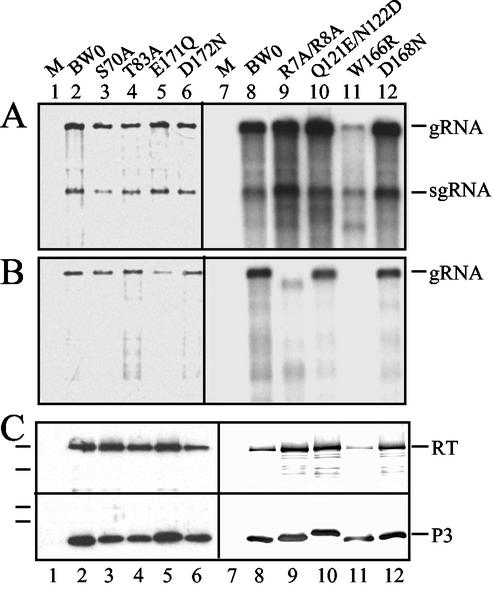
Point mutations were introduced into the major capsid protein (P3) of cloned infectious cDNA of the polerovirus beet western yellows virus (BWYV) by manipulation of cloned infectious cDNA. Seven mutations targeted sites on the S domain predicted to lie on the capsid surface. An eighth mutation eliminated two arginine residues in the R domain, which is thought to extend into the capsid interior. The effects of the mutations on virus capsid formation, virus accumulation in protoplasts and plants, and aphid transmission were tested. All of the mutants replicated in protoplasts. The S-domain mutant W166R failed to protect viral RNA from RNase attack, suggesting that this particular mutation interfered with stable capsid formation. The R-domain mutant R7A/R8A protected approximately 90% of the viral RNA strand from RNase, suggesting that lower positive-charge density in the mutant capsid interior interfered with stable packaging of the complete strand into virions. Neither of these mutants systemically infected plants. The six remaining mutants properly packaged viral RNA and could invade Nicotiana clevelandii systemically following agroinfection. Mutant Q121E/N122D was poorly transmitted by aphids, implicating one or both targeted residues in virus-vector interactions. Successful transmission of mutant D172N was accompanied either by reversion to the wild type or by appearance of a second-site mutation, N137D. This finding indicates that D172 is also important for transmission but that the D172N transmission defect can be compensated for by a "reverse" substitution at another site. The results have been used to evaluate possible structural models for the BWYV capsid.
Figures
References
-
- Brault, V., J. Mutterer, D. Scheidecker, M. T. Simonis, E. Herrbach, K. Richards, and V. Ziegler-Graff. 2000. Effects of point mutations in the readthrough domain of beet western yellows virus minor capsid protein on virus accumulation in planta and on transmission by aphids. J. Virol. 74:1140-1148. - PMC - PubMed
-
- Bruyère, A., V. Brault, V. Ziegler-Graff, M. T. Simonis, J. F. J. M. van den Heuvel, K. Richards, H. Guilley, G. Jonard, and E. Herrbach. 1997. Effects of mutations in the beet western yellows virus readthrough protein on its expression and packaging, and on virus accumulation, symptoms and aphid transmission. Virology 230:323-334. - PubMed
-
- Cheng, S. L., L. L. Domier, and C. J. D'Arcy. 1994. Detection of the readthrough protein of barley yellow dwarf virus. Virology 202:1003-1006. - PubMed
-
- Dolja, V. V., and E. V. Koonin. 1991. Phylogeny of capsid proteins of small icosahedral RNA plant viruses. J. Gen. Virol. 72:1481-1486. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
