

Subcellular targeting of methylmercury lyase enhances its specific activity for organic mercury detoxification in plants
- PMID: 12586871
- PMCID: PMC166823
- DOI: 10.1104/pp.010124
Subcellular targeting of methylmercury lyase enhances its specific activity for organic mercury detoxification in plants
Abstract
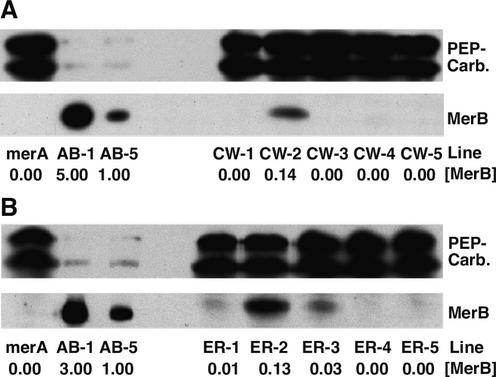
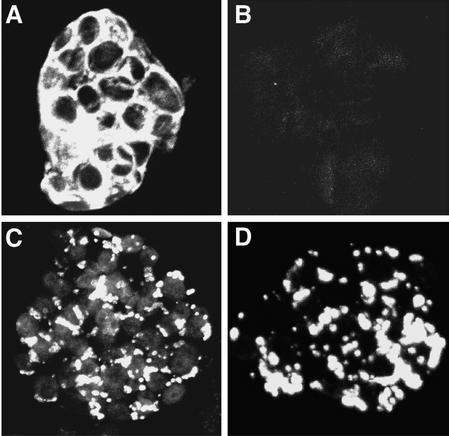
Methylmercury is an environmental pollutant that biomagnifies in the aquatic food chain with severe consequences for humans and other animals. In an effort to remove this toxin in situ, we have been engineering plants that express the bacterial mercury resistance enzymes organomercurial lyase MerB and mercuric ion reductase MerA. In vivo kinetics experiments suggest that the diffusion of hydrophobic organic mercury to MerB limits the rate of the coupled reaction with MerA (Bizily et al., 2000). To optimize reaction kinetics for organic mercury compounds, the merB gene was engineered to target MerB for accumulation in the endoplasmic reticulum and for secretion to the cell wall. Plants expressing the targeted MerB proteins and cytoplasmic MerA are highly resistant to organic mercury and degrade organic mercury at 10 to 70 times higher specific activity than plants with the cytoplasmically distributed wild-type MerB enzyme. MerB protein in endoplasmic reticulum-targeted plants appears to accumulate in large vesicular structures that can be visualized in immunolabeled plant cells. These results suggest that the toxic effects of organic mercury are focused in microenvironments of the secretory pathway, that these hydrophobic compartments provide more favorable reaction conditions for MerB activity, and that moderate increases in targeted MerB expression will lead to significant gains in detoxification. In summary, to maximize phytoremediation efficiency of hydrophobic pollutants in plants, it may be beneficial to target enzymes to specific subcellular environments.
Figures



Similar articles
-
Phytodetoxification of hazardous organomercurials by genetically engineered plants.Nat Biotechnol. 2000 Feb;18(2):213-7. doi: 10.1038/72678. Nat Biotechnol. 2000. PMID: 10657131
-
Organomercurial Lyase (MerB)-Mediated Demethylation Decreases Bacterial Methylmercury Resistance in the Absence of Mercuric Reductase (MerA).Appl Environ Microbiol. 2022 Mar 22;88(6):e0001022. doi: 10.1128/aem.00010-22. Epub 2022 Feb 9. Appl Environ Microbiol. 2022. PMID: 35138926 Free PMC article.
-
Development of a transgenic tobacco plant for phytoremediation of methylmercury pollution.Appl Microbiol Biotechnol. 2010 Jun;87(2):781-6. doi: 10.1007/s00253-010-2572-9. Appl Microbiol Biotechnol. 2010. PMID: 20393701
-
Strategies for the engineered phytoremediation of toxic element pollution: mercury and arsenic.J Ind Microbiol Biotechnol. 2005 Dec;32(11-12):502-13. doi: 10.1007/s10295-005-0255-9. Epub 2005 Jul 2. J Ind Microbiol Biotechnol. 2005. PMID: 15995854 Review.
-
Genetic engineering to enhance mercury phytoremediation.Curr Opin Biotechnol. 2009 Apr;20(2):213-9. doi: 10.1016/j.copbio.2009.02.010. Epub 2009 Mar 26. Curr Opin Biotechnol. 2009. PMID: 19328673 Free PMC article. Review.
Cited by
-
Phytoremediation and Microorganisms-Assisted Phytoremediation of Mercury-Contaminated Soils: Challenges and Perspectives.Int J Environ Res Public Health. 2021 Mar 2;18(5):2435. doi: 10.3390/ijerph18052435. Int J Environ Res Public Health. 2021. PMID: 33801363 Free PMC article. Review.
-
Long-distance root-to-shoot transport of phytochelatins and cadmium in Arabidopsis.Proc Natl Acad Sci U S A. 2003 Aug 19;100(17):10118-23. doi: 10.1073/pnas.1734072100. Epub 2003 Aug 8. Proc Natl Acad Sci U S A. 2003. PMID: 12909714 Free PMC article.
-
Transgenic merA and merB expression reduces mercury contamination in vegetables and grains grown in mercury-contaminated soil.Plant Cell Rep. 2020 Oct;39(10):1369-1380. doi: 10.1007/s00299-020-02570-8. Epub 2020 Jul 25. Plant Cell Rep. 2020. PMID: 32712731
-
A bacterial view of the periodic table: genes and proteins for toxic inorganic ions.J Ind Microbiol Biotechnol. 2005 Dec;32(11-12):587-605. doi: 10.1007/s10295-005-0019-6. Epub 2005 Oct 12. J Ind Microbiol Biotechnol. 2005. PMID: 16133099 Review.
-
SCARECROW promoter-driven expression of a bacterial mercury transporter MerC in root endodermal cells enhances mercury accumulation in Arabidopsis shoots.Planta. 2019 Aug;250(2):667-674. doi: 10.1007/s00425-019-03186-3. Epub 2019 May 18. Planta. 2019. PMID: 31104129
References
-
- Balogh SJ, Huang Y, Offerman HJ, Meyer ML, Johnson DK. Episodes of elevated methylmercury concentrations in prairie streams. Environ Sci Technol. 2002;36:1665–1670. - PubMed
-
- Begley TP, Walts AE, Walsh CT. Mechanistic studies of a protonolytic organomercurial cleaving enzyme: bacterial organomercurial lyase. Biochemistry. 1986;25:7192–7200. - PubMed
-
- Bizily S, Rugh CL, Meagher RB. Phytodetoxification of hazardous organomercurials by genetically engineered plants. Nat Biotechnol. 2000;18:213–217. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
