

Expression of a bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase in transgenic rice plants increases trehalose accumulation and abiotic stress tolerance without stunting growth
- PMID: 12586876
- PMCID: PMC166828
- DOI: 10.1104/pp.007237
Expression of a bifunctional fusion of the Escherichia coli genes for trehalose-6-phosphate synthase and trehalose-6-phosphate phosphatase in transgenic rice plants increases trehalose accumulation and abiotic stress tolerance without stunting growth
Abstract
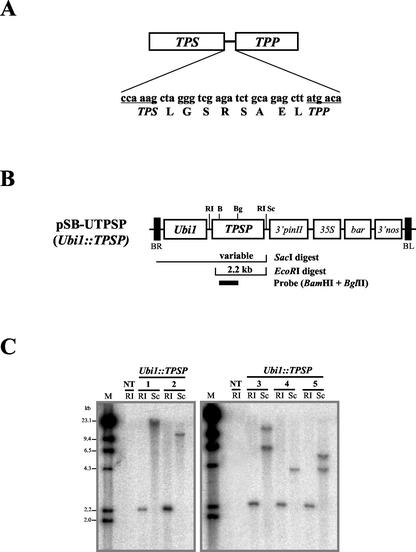
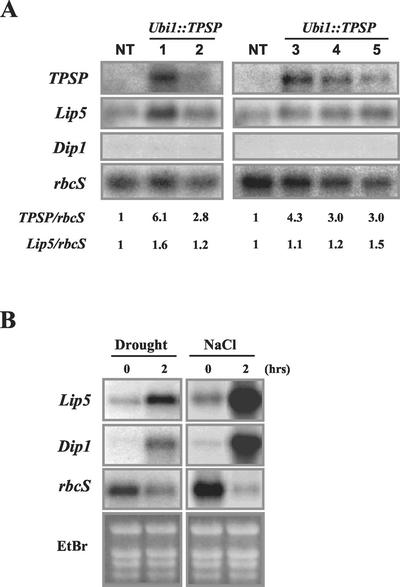
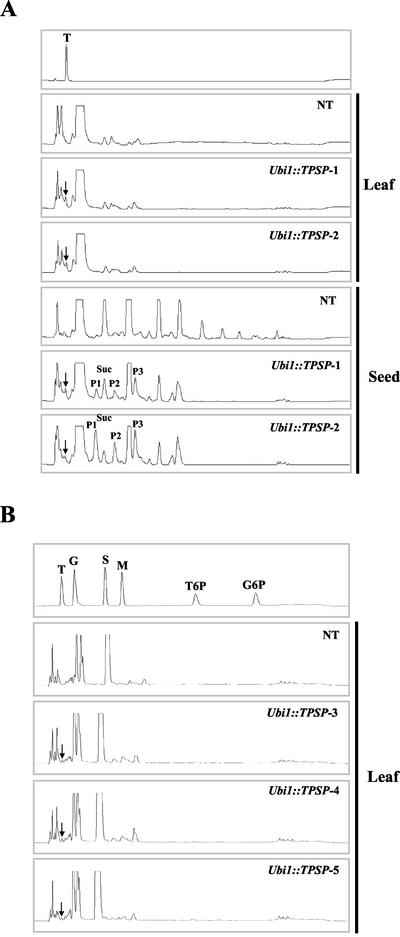
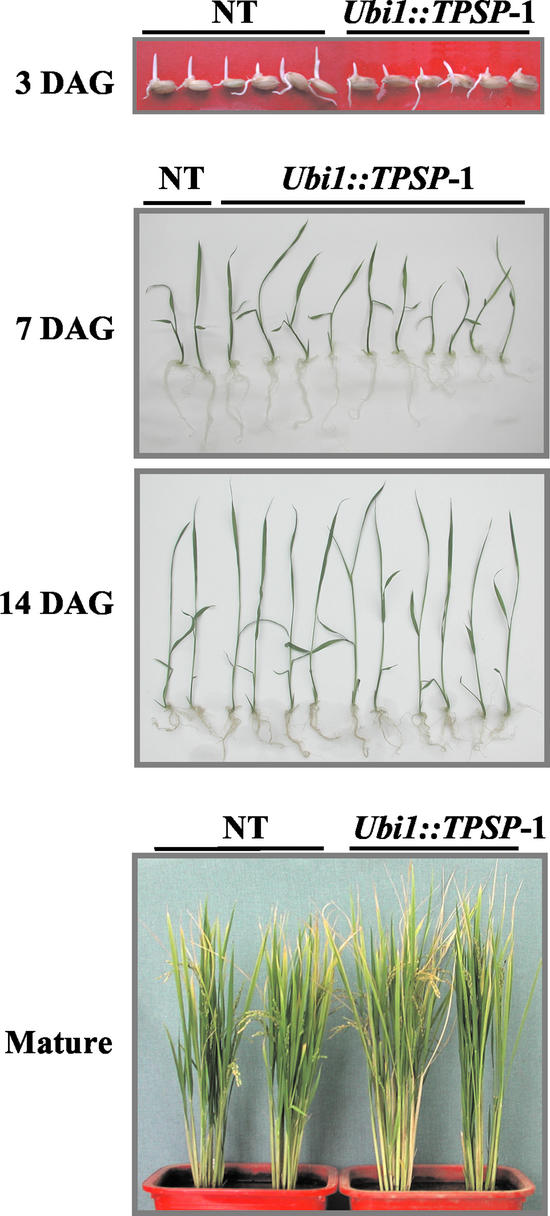
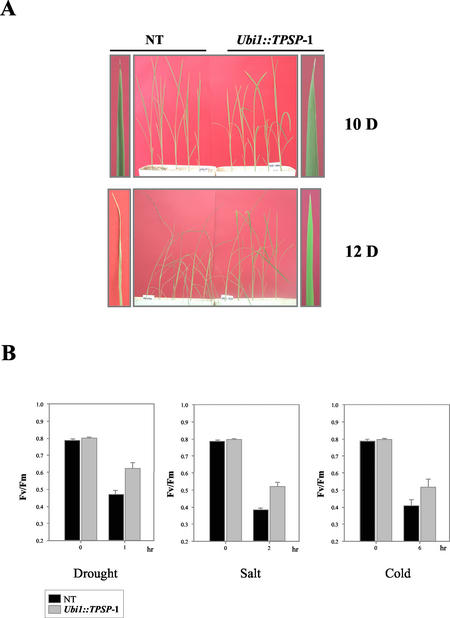
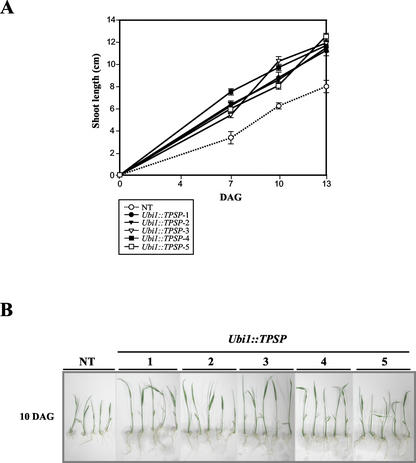
Trehalose plays an important role in stress tolerance in plants. Trehalose-producing, transgenic rice (Oryza sativa) plants were generated by the introduction of a gene encoding a bifunctional fusion (TPSP) of the trehalose-6-phosphate (T-6-P) synthase (TPS) and T-6-P phosphatase (TPP) of Escherichia coli, under the control of the maize (Zea mays) ubiquitin promoter (Ubi1). The high catalytic efficiency (Seo et al., 2000) of the fusion enzyme and the single-gene engineering strategy make this an attractive candidate for high-level production of trehalose; it has the added advantage of reducing the accumulation of potentially deleterious T-6-P. The trehalose levels in leaf and seed extracts from Ubi1::TPSP plants were increased up to 1.076 mg g fresh weight(-1). This level was 200-fold higher than that of transgenic tobacco (Nicotiana tabacum) plants transformed independently with either TPS or TPP expression cassettes. The carbohydrate profiles were significantly altered in the seeds, but not in the leaves, of Ubi1::TPSP plants. It has been reported that transgenic plants with E. coli TPS and/or TPP were severely stunted and root morphology was altered. Interestingly, our Ubi1::TPSP plants showed no growth inhibition or visible phenotypic alterations despite the high-level production of trehalose. Moreover, trehalose accumulation in Ubi1::TPSP plants resulted in increased tolerance to drought, salt, and cold, as shown by chlorophyll fluorescence and growth inhibition analyses. Thus, our results suggest that trehalose acts as a global protectant against abiotic stress, and that rice is more tolerant to trehalose synthesis than dicots.
Figures
Similar articles
-
Characterization of a bifunctional enzyme fusion of trehalose-6-phosphate synthetase and trehalose-6-phosphate phosphatase of Escherichia coli.Appl Environ Microbiol. 2000 Jun;66(6):2484-90. doi: 10.1128/AEM.66.6.2484-2490.2000. Appl Environ Microbiol. 2000. PMID: 10831428 Free PMC article.
-
A bifunctional TPS-TPP enzyme from yeast confers tolerance to multiple and extreme abiotic-stress conditions in transgenic Arabidopsis.Planta. 2007 Nov;226(6):1411-21. doi: 10.1007/s00425-007-0579-y. Epub 2007 Jul 13. Planta. 2007. PMID: 17628825
-
Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses.Proc Natl Acad Sci U S A. 2002 Dec 10;99(25):15898-903. doi: 10.1073/pnas.252637799. Epub 2002 Nov 27. Proc Natl Acad Sci U S A. 2002. PMID: 12456878 Free PMC article.
-
Trehalose biosynthesis in response to abiotic stresses.J Integr Plant Biol. 2008 Oct;50(10):1223-9. doi: 10.1111/j.1744-7909.2008.00736.x. J Integr Plant Biol. 2008. PMID: 19017109 Review.
-
Trehalose metabolism in Escherichia coli: stress protection and stress regulation of gene expression.Mol Microbiol. 1993 Apr;8(2):205-10. doi: 10.1111/j.1365-2958.1993.tb01564.x. Mol Microbiol. 1993. PMID: 8391102 Review.
Cited by
-
Genome-Wide Identification, Evolution, and Expression Analysis of TPS and TPP Gene Families in Brachypodium distachyon.Plants (Basel). 2019 Sep 23;8(10):362. doi: 10.3390/plants8100362. Plants (Basel). 2019. PMID: 31547557 Free PMC article.
-
Heterologous mannitol-1-phosphate dehydrogenase gene over-expression in Parachlorella kessleri for enhanced microalgal biomass productivity.J Genet Eng Biotechnol. 2022 Feb 28;20(1):38. doi: 10.1186/s43141-022-00322-7. J Genet Eng Biotechnol. 2022. PMID: 35226194 Free PMC article.
-
Comparative transcriptome analysis reveals molecular regulation of salt tolerance in two contrasting chickpea genotypes.Front Plant Sci. 2023 May 30;14:1191457. doi: 10.3389/fpls.2023.1191457. eCollection 2023. Front Plant Sci. 2023. PMID: 37360702 Free PMC article.
-
A novel TF molecular switch-mechanism found in two contrasting ecotypes of a psammophyte, Agriophyllum squarrosum, in regulating transcriptional drought memory.BMC Plant Biol. 2023 Mar 30;23(1):167. doi: 10.1186/s12870-023-04154-6. BMC Plant Biol. 2023. PMID: 36997861 Free PMC article.
-
Enhancing Salt Tolerance of Plants: From Metabolic Reprogramming to Exogenous Chemical Treatments and Molecular Approaches.Cells. 2020 Nov 17;9(11):2492. doi: 10.3390/cells9112492. Cells. 2020. PMID: 33212751 Free PMC article. Review.
References
-
- Aguan K, Sugawara K, Suzuki N, Kusano T. Isolation of genes for low-temperature-induced proteins in rice by a simple subtractive method. Plant Cell Physiol. 1991;32:1285–1289.
-
- Blazquez MA, Lagunas R, Gancedo C, Gancedo JM. Trehalose-6-phosphate, a new regulator of yeast glycolysis that inhibits hexokinases. FEBS Lett. 1993;329:51–54. - PubMed
-
- Bradford M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
