

Reciprocal and nonreciprocal recombination at the glucocerebrosidase gene region: implications for complexity in Gaucher disease
- PMID: 12587096
- PMCID: PMC1180228
- DOI: 10.1086/367850
Reciprocal and nonreciprocal recombination at the glucocerebrosidase gene region: implications for complexity in Gaucher disease
Abstract
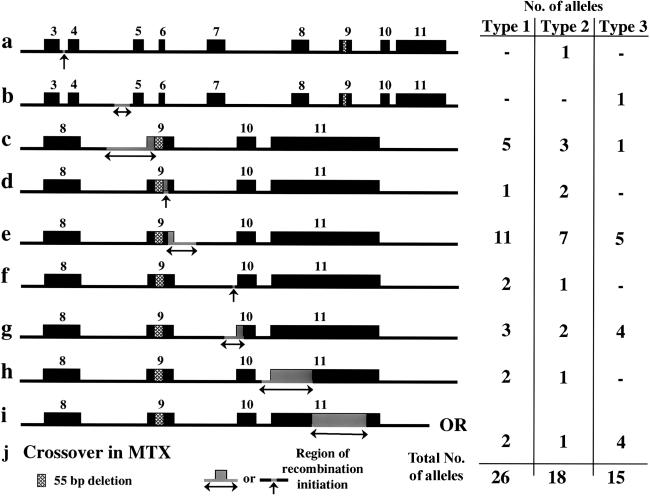
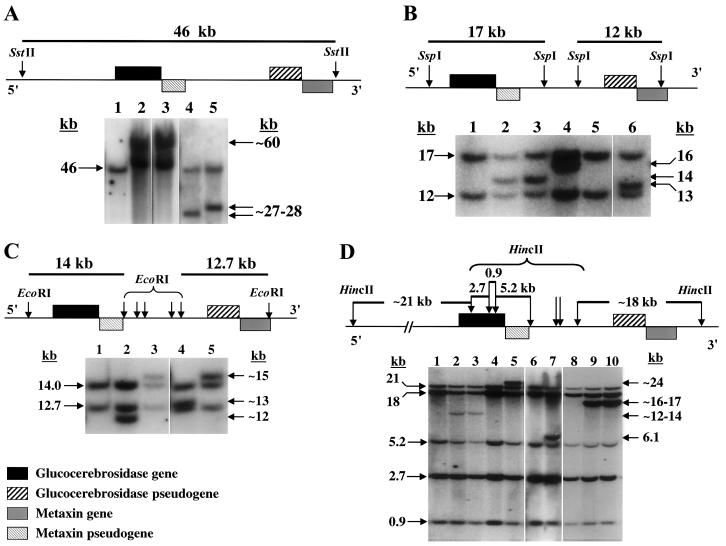
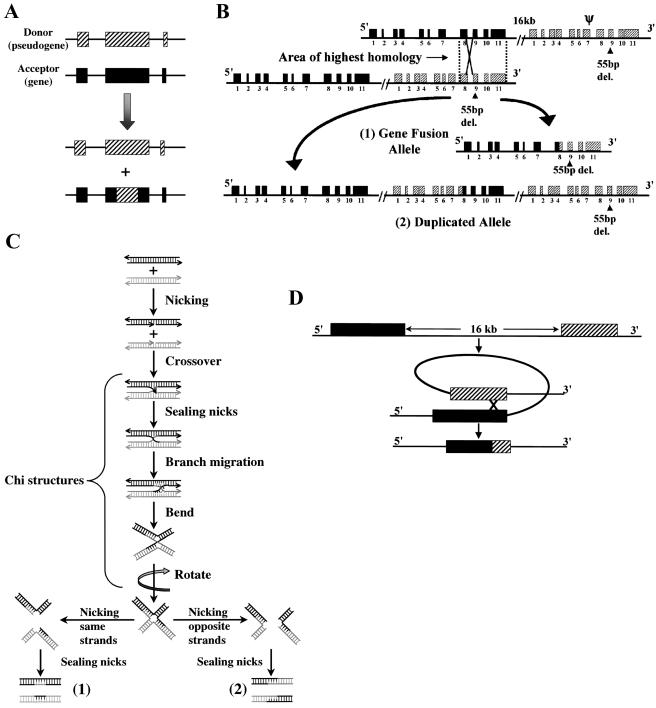
Gaucher disease results from an autosomal recessive deficiency of the lysosomal enzyme glucocerebrosidase. The glucocerebrosidase gene is located in a gene-rich region of 1q21 that contains six genes and two pseudogenes within 75 kb. The presence of contiguous, highly homologous pseudogenes for both glucocerebrosidase and metaxin at the locus increases the likelihood of DNA rearrangements in this region. These recombinations can complicate genotyping in patients with Gaucher disease and contribute to the difficulty in interpreting genotype-phenotype correlations in this disorder. In the present study, DNA samples from 240 patients with Gaucher disease were examined using several complementary approaches to identify and characterize recombinant alleles, including direct sequencing, long-template polymerase chain reaction, polymorphic microsatellite repeats, and Southern blots. Among the 480 alleles studied, 59 recombinant alleles were identified, including 34 gene conversions, 18 fusions, and 7 downstream duplications. Twenty-two percent of the patients evaluated had at least one recombinant allele. Twenty-six recombinant alleles were found among 310 alleles from patients with type 1 disease, 18 among 74 alleles from patients with type 2 disease, and 15 among 96 alleles from patients with type 3 disease. Several patients carried two recombinations or mutations on the same allele. Generally, alleles resulting from nonreciprocal recombination (gene conversion) could be distinguished from those arising by reciprocal recombination (crossover and exchange), and the length of the converted sequence was determined. Homozygosity for a recombinant allele was associated with early lethality. Ten different sites of crossover and a shared pentamer motif sequence (CACCA) that could be a hotspot for recombination were identified. These findings contribute to a better understanding of genotype-phenotype relationships in Gaucher disease and may provide insights into the mechanisms of DNA rearrangement in other disorders.
Figures
References
Electronic-Database Information
-
- GenBank, http://www.ncbi.nlm.nih.gov/Genbank/ (for human GBA [accession number J03059], the human GBA pseudogene [accession number J03060], and the sequences of genes surrounding human GBA [accession number AF023263])
-
- Human Genetic Disease Databank (GeneDis), http://life2.tau.ac.il/GeneDis/ (for compilation of mutations in human glucocerebrosidase)
-
- Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/Omim/ (for Gaucher disease [MIM 230800], Gaucher disease type 2 [MIM 230900], and Gaucher disease type 3 [MIM 231000])
References
-
- Aarskog NK, Vedeler CA (2001) Recombination breakpoints in the Charcot-Marie-Tooth 1A repeat sequence in Norwegian families. Acta Neurol Scand 104:97–100 - PubMed
-
- Armstrong LC, Komiya T, Bergman BE, Mihara K, Bornstein P (1997) Metaxin is a component of a preprotein import complex in the outer membrane of the mammalian mitochondrion. J Biol Chem 272:6510–6518 - PubMed
-
- Batzer MA, Deininger PL (2002) Alu repeats and human genomic diversity. Nat Rev Genet 3:370–379 - PubMed
-
- Bell SJ, Chow YC, Ho JY, Forsdyke DR (1998) Correlation of chi orientation with transcription indicates a fundamental relationship between recombination and transcription. Gene 216:285–292 - PubMed
-
- Beutler E, Grabowski GA (2001) Gaucher disease. In: Scriver CR, Beaudet AL, Valle D, Sly WS (eds) The metabolic and molecular bases of inherited disease. McGraw-Hill, New York, pp 3635–3668
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
