

The role of prolactin in mammary carcinoma
- PMID: 12588805
- PMCID: PMC1698952
- DOI: 10.1210/er.2001-0036
The role of prolactin in mammary carcinoma
Abstract
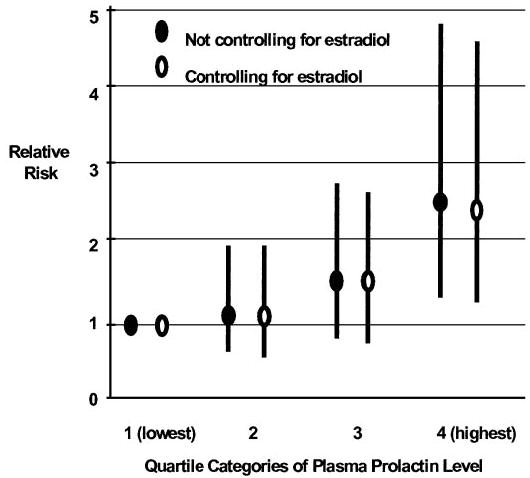
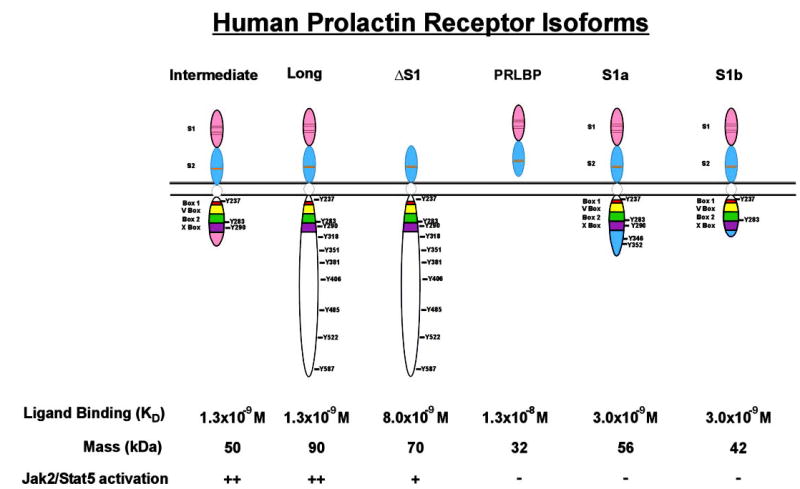
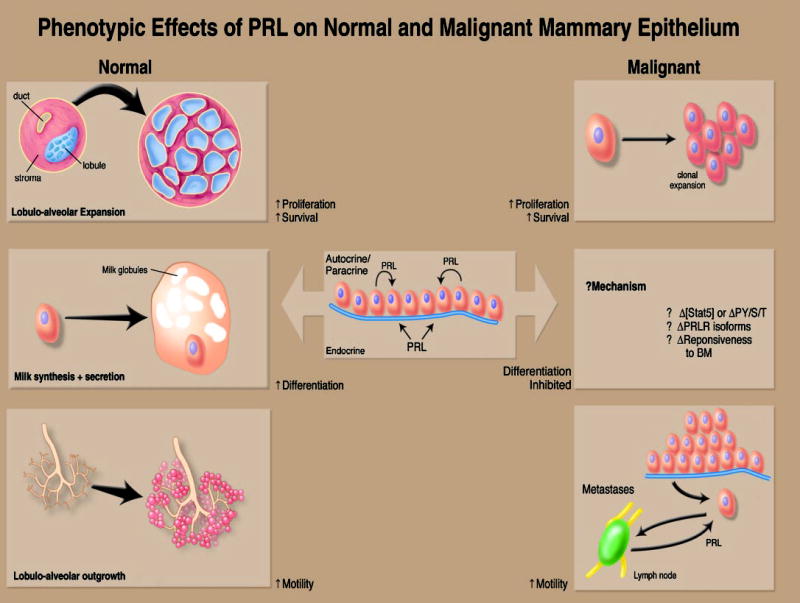
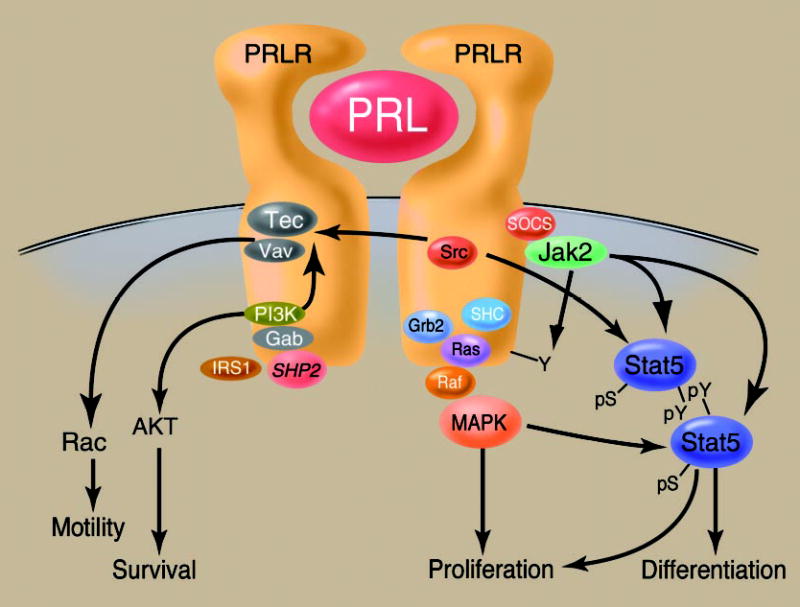
The contribution of prolactin (PRL) to the pathogenesis and progression of human breast cancer at the cellular, transgenic, and epidemiological levels is increasingly appreciated. Acting at the endocrine and autocrine/paracrine levels, PRL functions to stimulate the growth and motility of human breast cancer cells. The actions of this ligand are mediated by at least six recognized PRL receptor isoforms found on, or secreted by, human breast epithelium. The PRL/PRL receptor complex associates with and activates several signaling networks that are shared with other members of the cytokine receptor superfamily. Coupled with the recently identified intranuclear function of PRL, these networks are integrated into the in vitro and in vivo actions induced by ligand. These findings indicate that antagonists of PRL/PRL receptor interaction or PRL receptor-associated signal transduction may be of considerable utility in the treatment of human breast cancer.
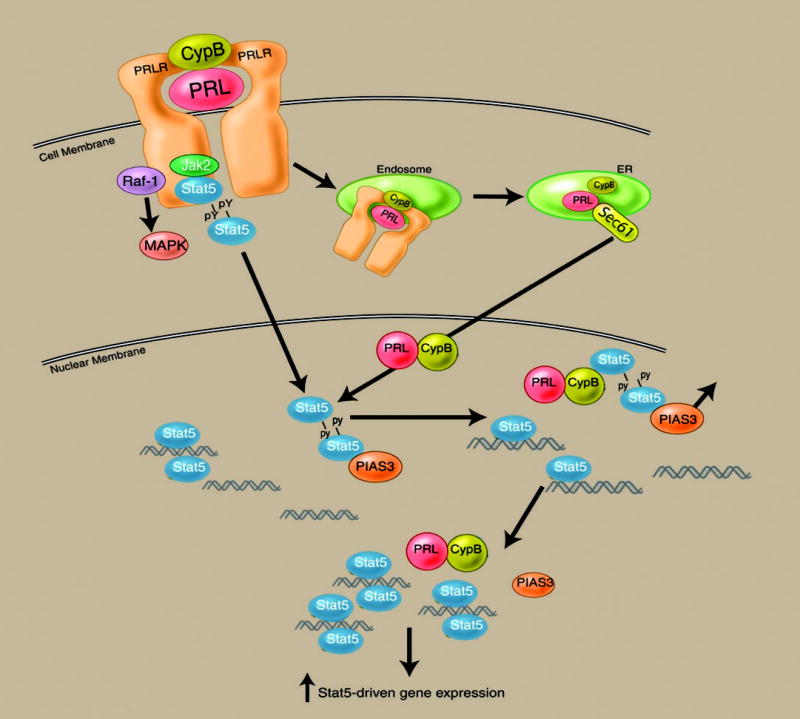
Figures
References
-
- Welsch CW, Nagasawa H. Prolactin and murine mammary tumorigenesis: a review. Cancer Res. 1977;37:951–963. - PubMed
-
- Dickson RB, Johnson MD, Bano M, Shi E, Kurebayashi J, Ziff B, Martinez-Lacaci I, Amundadottir LT, Lippman ME. Growth factors in breast cancer: mitogenesis to transformation. J Steroid Biochem Mol Biol. 1992;43:69 –78. - PubMed
-
- Love RR, Rose DR, Surawicz TS, Newcomb PA. Prolactin and growth hormone levels in premenopausal women with breast cancer and healthy women with a strong family history of breast cancer. Cancer. 1991;68:1401–1405. - PubMed
-
- Kwa HG, Wang DY. An abnormal luteal-phase evening peak of plasma prolactin in women with a family history of breast cancer. Int J Cancer. 1977;20:12–14. - PubMed
-
- Musey VC, Collins DC, Musey PI, Martino-Saltzman D, Preedy JR. Long-term effect of a first pregnancy on the secretion of prolactin. N Engl J Med. 1987;316:229 –234. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- CA-49449/CA/NCI NIH HHS/United States
- P01 CA087969/CA/NCI NIH HHS/United States
- R01 CA067262/CA/NCI NIH HHS/United States
- CA-87969/CA/NCI NIH HHS/United States
- U01 CA067262/CA/NCI NIH HHS/United States
- DK-92265/DK/NIDDK NIH HHS/United States
- T32 HD007259/HD/NICHD NIH HHS/United States
- R01 CA069294/CA/NCI NIH HHS/United States
- CA-8312/CA/NCI NIH HHS/United States
- CA-67262/CA/NCI NIH HHS/United States
- T32 HD-07259/HD/NICHD NIH HHS/United States
- R01 CA049449/CA/NCI NIH HHS/United States
- R01 CA078312/CA/NCI NIH HHS/United States
- U01 CA049449/CA/NCI NIH HHS/United States
- R01 CA-69294/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
