

Inhibitory and stimulatory regulation of Rac and cell motility by the G12/13-Rho and Gi pathways integrated downstream of a single G protein-coupled sphingosine-1-phosphate receptor isoform
- PMID: 12588974
- PMCID: PMC151702
- DOI: 10.1128/MCB.23.5.1534-1545.2003
Inhibitory and stimulatory regulation of Rac and cell motility by the G12/13-Rho and Gi pathways integrated downstream of a single G protein-coupled sphingosine-1-phosphate receptor isoform
Abstract
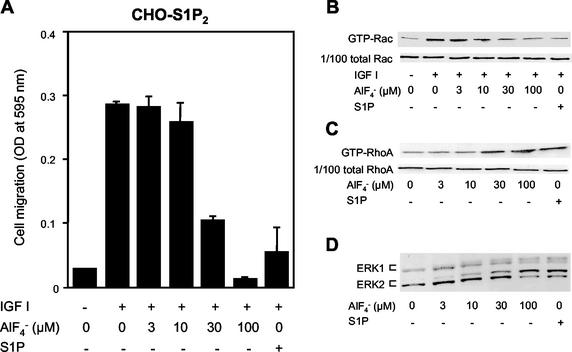
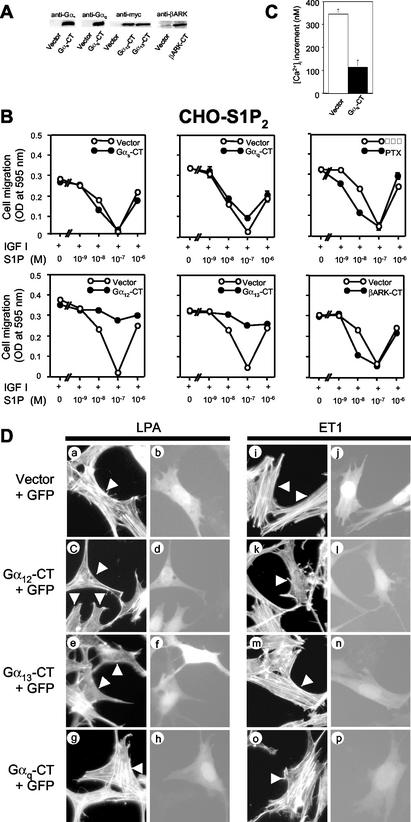
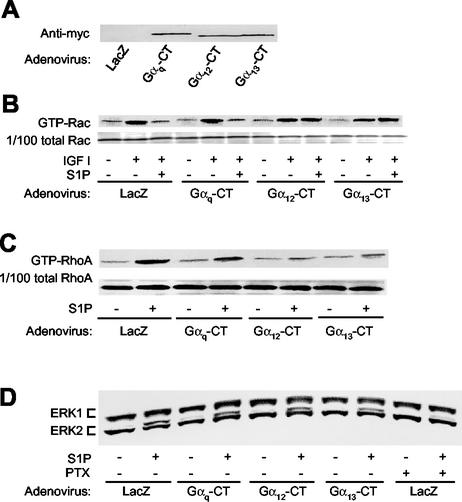
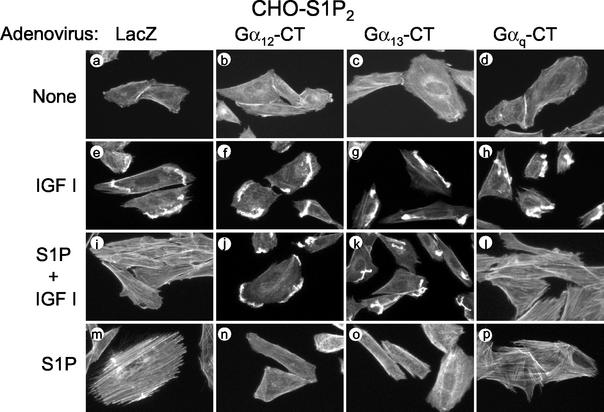
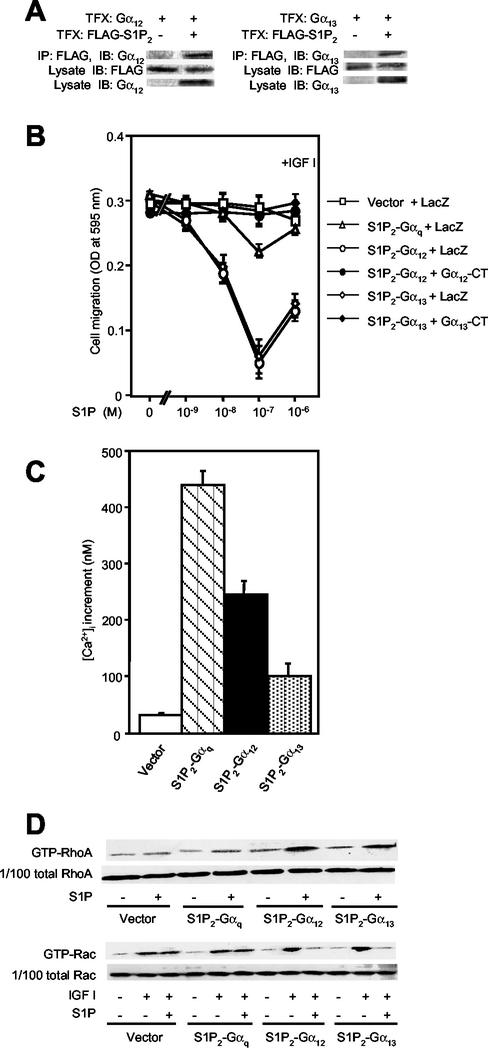
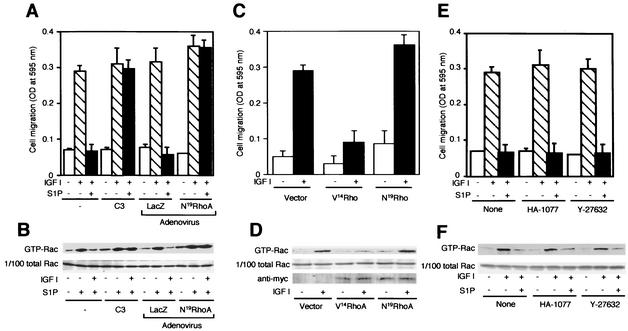
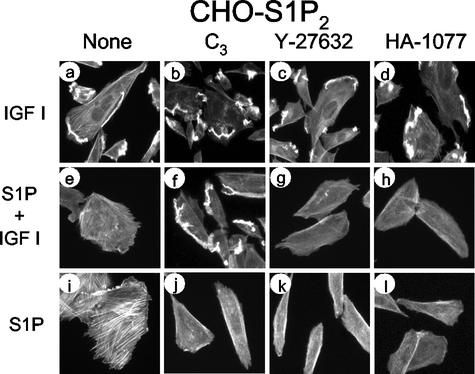
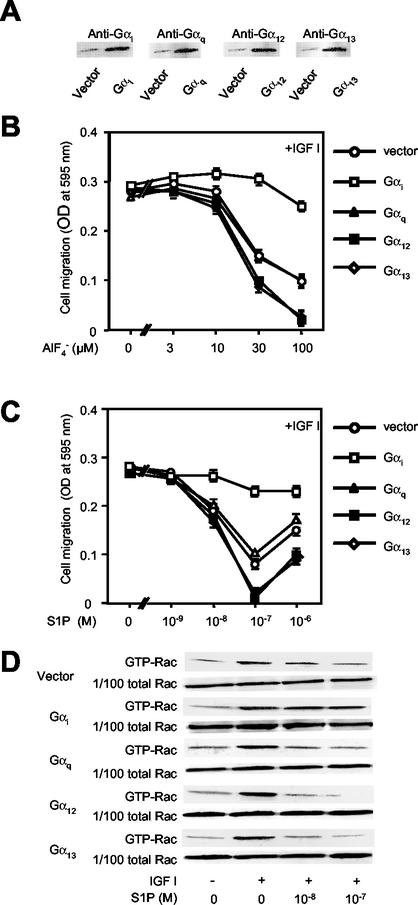
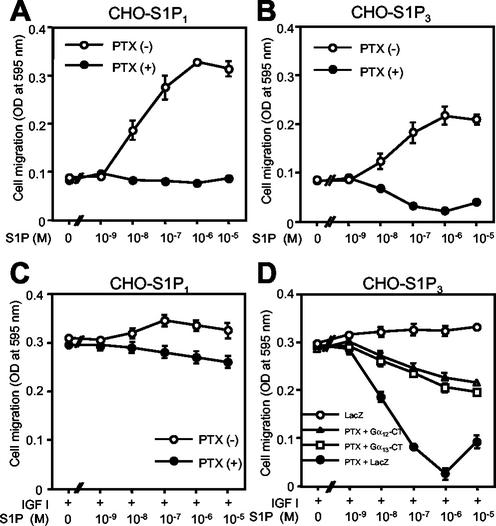
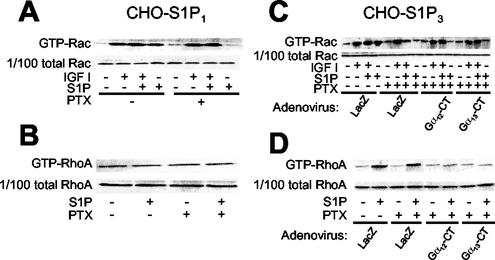
The G protein-coupled receptors S1P2/Edg5 and S1P3/Edg3 both mediate sphingosine-1-phosphate (S1P) stimulation of Rho, yet S1P2 but not S1P3 mediates downregulation of Rac activation, membrane ruffling, and cell migration in response to chemoattractants. Specific inhibition of endogenous Galpha12 and Galpha13, but not of Galphaq, by expression of respective C-terminal peptides abolished S1P2-mediated inhibition of Rac, membrane ruffling, and migration, as well as stimulation of Rho and stress fiber formation. Fusion receptors comprising S1P2 and either Galpha12 or Galpha13, but not Galphaq, mediated S1P stimulation of Rho and also inhibition of Rac and migration. Overexpression of Galphai, by contrast, specifically antagonized S1P2-mediated inhibition of Rac and migration. The S1P2 actions were mimicked by expression of V14Rho and were abolished by C3 toxin and N19Rho, but not Rho kinase inhibitors. In contrast to S1P2, S1P3 mediated S1P-directed, pertussis toxin-sensitive chemotaxis and Rac activation despite concurrent stimulation of Rho via G12/13. Upon inactivation of Gi by pertussis toxin, S1P3 mediated inhibition of Rac and migration just like S1P2. These results indicate that integration of counteracting signals from the Gi- and the G12/13-Rho pathways directs either positive or negative regulation of Rac, and thus cell migration, upon activation of a single S1P receptor isoform.
Figures
References
-
- Akhter, S. A., L. M. Luttrell, H. A. Rockman, G. Iaccarino, R. J. Lefkowitz, and W. J. Koch. 1998. Targeting the receptor-Gq interface to inhibit in vivo pressure overload myocardial hypertrophy. Science 280:574-577. - PubMed
-
- Banyard, J., B. Anand-Apte, M. Symons, and B. R. Zetter. 2000. Motility and invasion are differentially modulated by Rho family GTPases. Oncogene 19:580-591. - PubMed
-
- Bornfeldt, K. E., L. M. Graves, E. W. Raines, Y. Igarashi, G. Wayman, S. Yamamura, Y. Yatomi, J. S. Sidhu, E. G. Krebs, S. Hakomori, and R. Ross. 1995. Sphingosine-1-phosphate inhibits PDGF-induced chemotaxis of human arterial smooth muscle cells: spatial and temporal modulation of PDGF chemotactic signal transduction. J. Cell Biol. 130:193-206. - PMC - PubMed
-
- Brinkmann, V., M. D. Davis, C. E. Heise, R. Albert, S. Cottens, R. Hof, C. Bruns, E. Prieschl, T. Baumruker, P. Hiestand, C. A. Foster, M. Zollinger, and K. R. Lynch. 2002. The immune modulator FTY720 targets sphingosine 1-phosphate receptors. J. Biol. Chem. 277:21453-21457. - PubMed
-
- Brzostowski, J. A., and A. R. Kimmel. 2001. Signaling at zero G: G-protein-independent functions for 7-TM receptors. Trends Biochem. Sci. 26:291-297. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous