

Actin filament polymerization regulates gliding motility by apicomplexan parasites
- PMID: 12589042
- PMCID: PMC149980
- DOI: 10.1091/mbc.e02-08-0458
Actin filament polymerization regulates gliding motility by apicomplexan parasites
Abstract
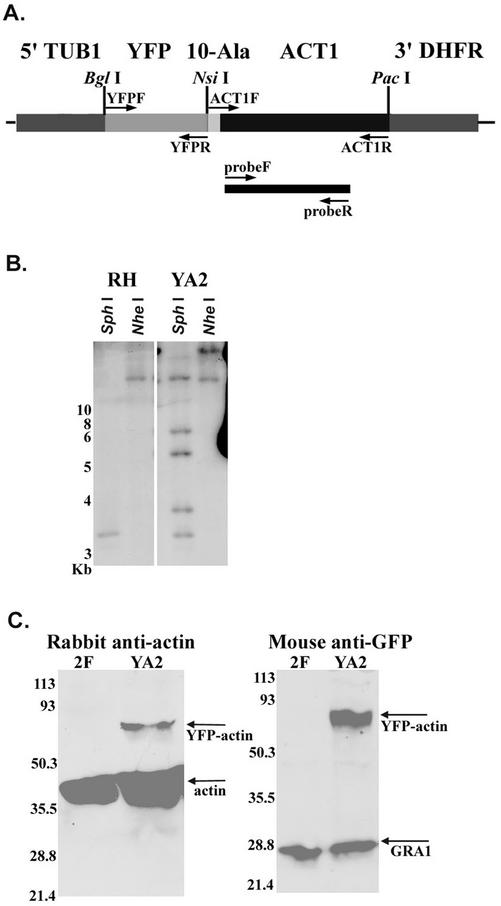
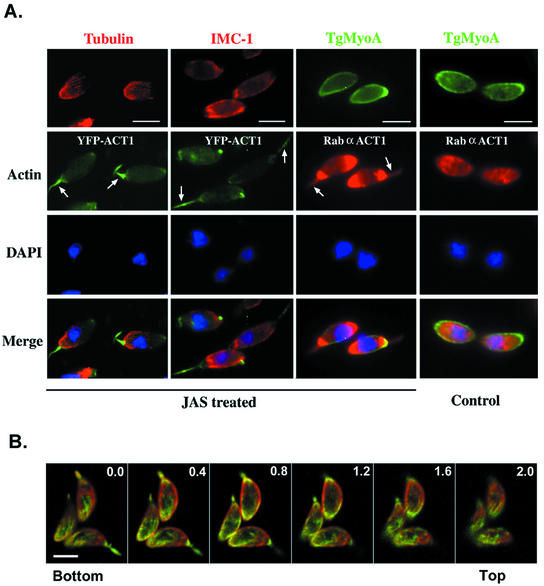
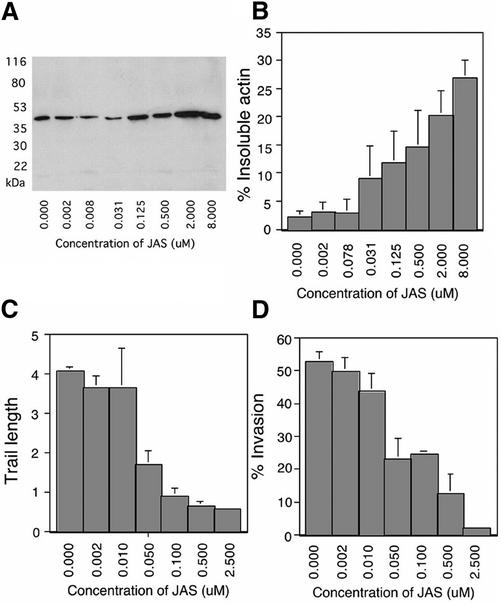
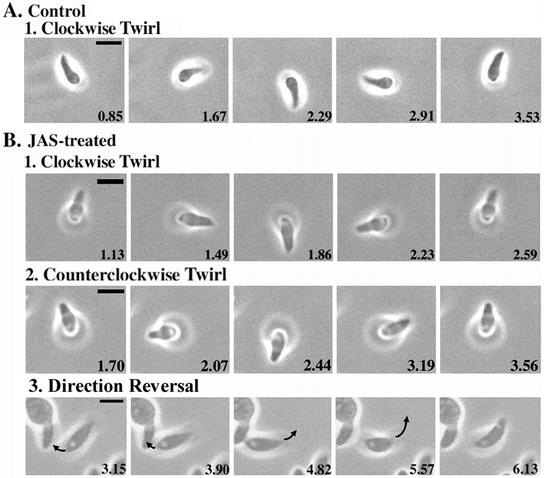
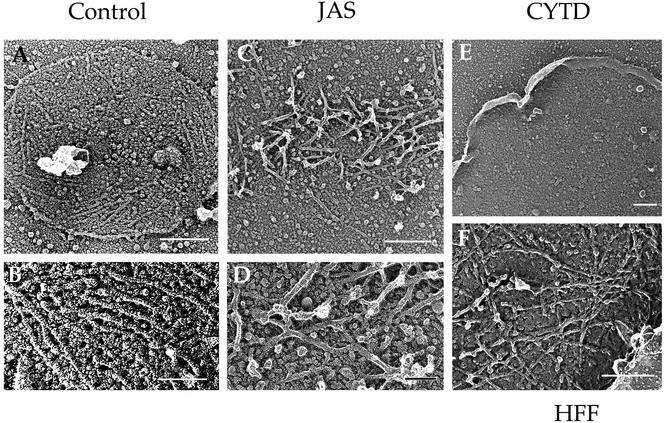
Host cell entry by Toxoplasma gondii depends critically on actin filaments in the parasite, yet paradoxically, its actin is almost exclusively monomeric. In contrast to the absence of stable filaments in conventional samples, rapid-freeze electron microscopy revealed that actin filaments were formed beneath the plasma membrane of gliding parasites. To investigate the role of actin filaments in motility, we treated parasites with the filament-stabilizing drug jasplakinolide (JAS) and monitored the distribution of actin in live and fixed cells using yellow fluorescent protein (YFP)-actin. JAS treatment caused YFP-actin to redistribute to the apical and posterior ends, where filaments formed a spiral pattern subtending the plasma membrane. Although previous studies have suggested that JAS induces rigor, videomicroscopy demonstrated that JAS treatment increased the rate of parasite gliding by approximately threefold, indicating that filaments are rate limiting for motility. However, JAS also frequently reversed the normal direction of motility, disrupting forward migration and cell entry. Consistent with this alteration, subcortical filaments in JAS-treated parasites occurred in tangled plaques as opposed to the straight, roughly parallel orientation observed in control cells. These studies reveal that precisely controlled polymerization of actin filaments imparts the correct timing, duration, and directionality of gliding motility in the Apicomplexa.
Figures
References
-
- Allen LH, Dobrowolski JM, Muller H, Sibley LD, Mansour TE. Cloning and characterization of actin depolymerizing factor from Toxoplasma gondii. Mol Biochem Parasitol. 1997;88:43–52. - PubMed
-
- Arrowood MJ, Sterling CR, Healey MC. Immunofluorescent microscopical visualization of trails left by gliding Cryptosporidium parvum sporozoites. J Parasitol. 1991;77:315–317. - PubMed
-
- Bubb MR, Senderowicz AMJ, Sausville EA, Duncan KLK, Korn ED. Jasplakinolide, a cytotoxic natural product, induces actin polymerization and completely inhibits the binding of phalloidin to F-actin. J Biol Chem. 1994;269:14869–14871. - PubMed
-
- Bubb MR, Spector I, Beyer BB, Fosen KM. Effects of jasplakinolide on the kinetics of actin polymerization. J Biol Chem. 2000;275:5163–5170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
