

Convergence of non-clathrin- and clathrin-derived endosomes involves Arf6 inactivation and changes in phosphoinositides
- PMID: 12589044
- PMCID: PMC149982
- DOI: 10.1091/mbc.02-04-0053
Convergence of non-clathrin- and clathrin-derived endosomes involves Arf6 inactivation and changes in phosphoinositides
Abstract
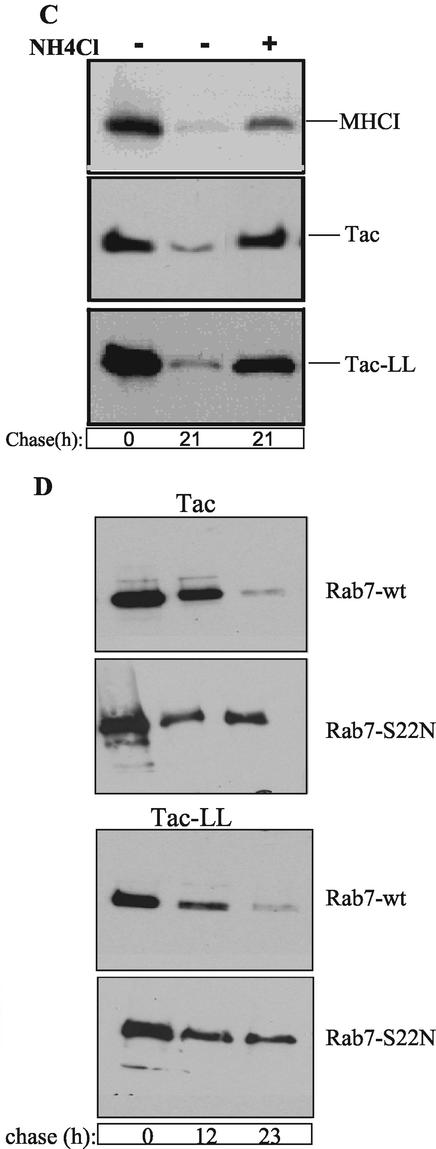
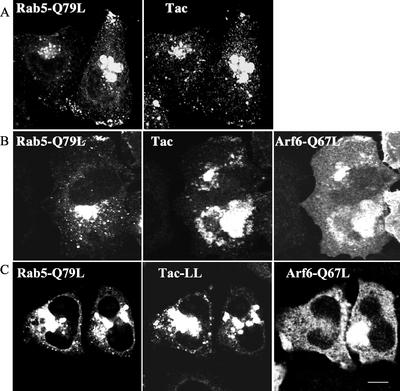
The trafficking of two plasma membrane (PM) proteins that lack clathrin internalization sequences, major histocompatibility complex class I (MHCI), and interleukin 2 receptor alpha subunit (Tac) was compared with that of PM proteins internalized via clathrin. MHCI and Tac were internalized into endosomes that were distinct from those containing clathrin cargo. At later times, a fraction of these internalized membranes were observed in Arf6-associated, tubular recycling endosomes whereas another fraction acquired early endosomal autoantigen 1 (EEA1) before fusion with the "classical" early endosomes containing the clathrin-dependent cargo, LDL. After convergence, cargo molecules from both pathways eventually arrived, in a Rab7-dependent manner, at late endosomes and were degraded. Expression of a constitutively active mutant of Arf6, Q67L, caused MHCI and Tac to accumulate in enlarged PIP(2)-enriched vacuoles, devoid of EEA1 and inhibited their fusion with clathrin cargo-containing endosomes and hence blocked degradation. By contrast, trafficking and degradation of clathrin-cargo was not affected. A similar block in transport of MHCI and Tac was reversibly induced by a PI3-kinase inhibitor, implying that inactivation of Arf6 and acquisition of PI3P are required for convergence of endosomes arising from these two pathways.
Figures







References
-
- Brown D, Rose J. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell. 1992;68:533–544. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
