

Involvement of the Arp2/3 complex and Scar2 in Golgi polarity in scratch wound models
- PMID: 12589062
- PMCID: PMC150000
- DOI: 10.1091/mbc.e02-06-0345
Involvement of the Arp2/3 complex and Scar2 in Golgi polarity in scratch wound models
Abstract
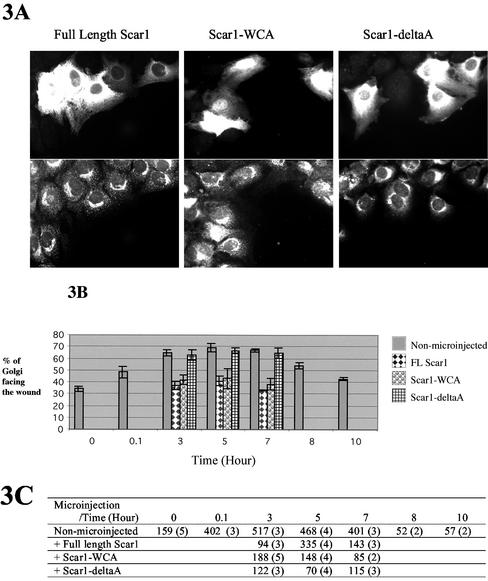
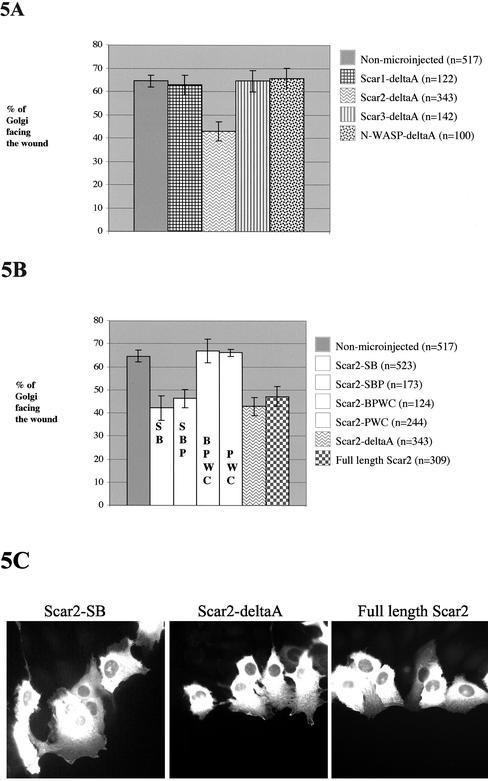
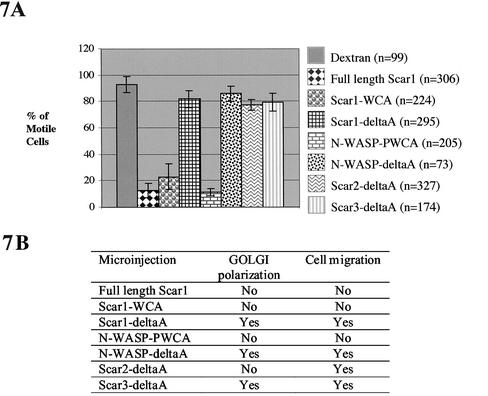
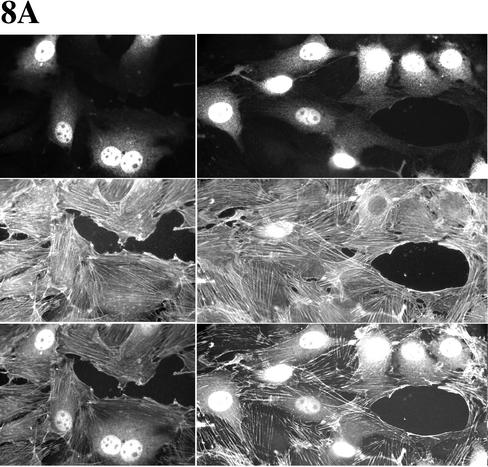
Cell motility and cell polarity are essential for morphogenesis, immune system function, and tissue repair. Many animal cells move by crawling, and one main driving force for movement is derived from the coordinated assembly and disassembly of actin filaments. As tissue culture cells migrate to close a scratch wound, this directional extension is accompanied by Golgi apparatus reorientation, to face the leading wound edge, giving the motile cell inherent polarity aligned relative to the wound edge and to the direction of cell migration. Cellular proteins essential for actin polymerization downstream of Rho family GTPases include the Arp2/3 complex as an actin nucleator and members of the Wiskott-Aldrich Syndrome protein (WASP) family as activators of the Arp2/3 complex. We therefore analyzed the involvement of the Arp2/3 complex and WASP-family proteins in in vitro wound healing assays using NIH 3T3 fibroblasts and astrocytes. In NIH 3T3 cells, we found that actin and Arp2/3 complex contributed to cell polarity establishment. Moreover, overexpression of N-terminal fragments of Scar2 (but not N-WASP or Scar1 or Scar3) interfere with NIH 3T3 Golgi polarization but not with cell migration. In contrast, actin, Arp2/3, and WASP-family proteins did not appear to be involved in Golgi polarization in astrocytes. Our results thus indicate that the requirement for Golgi polarity establishment is cell-type specific. Furthermore, in NIH 3T3 cells, Scar2 and the Arp2/3 complex appear to be involved in the establishment and maintenance of Golgi polarity during directed migration.
Figures








Similar articles
-
Microtubule involvement in NIH 3T3 Golgi and MTOC polarity establishment.J Cell Sci. 2003 Feb 15;116(Pt 4):743-56. doi: 10.1242/jcs.00288. J Cell Sci. 2003. PMID: 12538774
-
Scar1 and the related Wiskott-Aldrich syndrome protein, WASP, regulate the actin cytoskeleton through the Arp2/3 complex.Curr Biol. 1998 Dec 17-31;8(25):1347-56. doi: 10.1016/s0960-9822(98)00015-3. Curr Biol. 1998. PMID: 9889097
-
Different WASP family proteins stimulate different Arp2/3 complex-dependent actin-nucleating activities.Curr Biol. 2001 Dec 11;11(24):1903-13. doi: 10.1016/s0960-9822(01)00603-0. Curr Biol. 2001. PMID: 11747816
-
Regulation of actin filament network formation through ARP2/3 complex: activation by a diverse array of proteins.Annu Rev Biochem. 2001;70:649-76. doi: 10.1146/annurev.biochem.70.1.649. Annu Rev Biochem. 2001. PMID: 11395419 Review.
-
WASp in immune-system organization and function.Nat Rev Immunol. 2002 Sep;2(9):635-46. doi: 10.1038/nri884. Nat Rev Immunol. 2002. PMID: 12209132 Review.
Cited by
-
Targeting Cx43 and N-cadherin, which are abnormally upregulated in venous leg ulcers, influences migration, adhesion and activation of Rho GTPases.PLoS One. 2012;7(5):e37374. doi: 10.1371/journal.pone.0037374. Epub 2012 May 15. PLoS One. 2012. PMID: 22615994 Free PMC article.
-
Current methods for studying metastatic potential of tumor cells.Cancer Cell Int. 2022 Dec 9;22(1):394. doi: 10.1186/s12935-022-02801-w. Cancer Cell Int. 2022. PMID: 36494720 Free PMC article. Review.
-
Golgi polarization in a strong electric field.J Cell Sci. 2005 Mar 15;118(Pt 6):1117-28. doi: 10.1242/jcs.01646. Epub 2005 Feb 22. J Cell Sci. 2005. PMID: 15728257 Free PMC article.
-
Multifaceted Rho GTPase Signaling at the Endomembranes.Front Cell Dev Biol. 2019 Jul 16;7:127. doi: 10.3389/fcell.2019.00127. eCollection 2019. Front Cell Dev Biol. 2019. PMID: 31380367 Free PMC article. Review.
-
Correlation between liver metastasis of the colocalization of actin-related protein 2 and 3 complex and WAVE2 in colorectal carcinoma.Cancer Sci. 2007 Jul;98(7):992-9. doi: 10.1111/j.1349-7006.2007.00488.x. Epub 2007 Apr 23. Cancer Sci. 2007. PMID: 17459058 Free PMC article.
References
-
- Abercrombie M, Heaysman JE, Pegrum SM. The locomotion of fibroblasts in culture. I. Movements of the leading edge. Exp Cell Res. 1970;59:393–398. - PubMed
-
- Alrutz MA, Srivastava A, Wong KW, D'Souza-Schorey C, Tang M, Ch'Ng LE, Snapper SB, Isberg RR. Efficient uptake of Yersinia pseudotuberculosis via integrin receptors involves a Rac1-Arp 2/3 pathway that bypasses N-WASP function. Mol Microbiol. 2001;42:689–703. - PubMed
-
- Bailly M, Ichetovkin I, Grant W, Zebda N, Machesky LM, Segall JE, Condeelis J. The F-actin side binding activity of the Arp2/3 complex is essential for actin nucleation and lamellipod extension. Curr Biol. 2001;11:620–625. - PubMed
-
- Burns S, Thrasher AJ, Blundell MP, Machesky L, Jones GE. Configuration of human dendritic cell cytoskeleton by Rho GTPases, the WAS protein, and differentiation. Blood. 2001;98:1142–1149. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
