

Viable nonsense mutants for the essential gene SUP45 of Saccharomyces cerevisiae
- PMID: 12589713
- PMCID: PMC150568
- DOI: 10.1186/1471-2199-4-2
Viable nonsense mutants for the essential gene SUP45 of Saccharomyces cerevisiae
Abstract
Background: Termination of protein synthesis in eukaryotes involves at least two polypeptide release factors (eRFs) - eRF1 and eRF3. The highly conserved translation termination factor eRF1 in Saccharomyces cerevisiae is encoded by the essential gene SUP45.
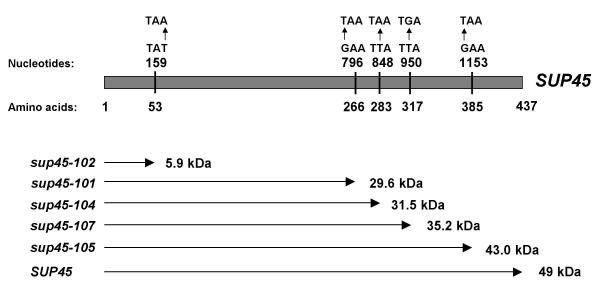
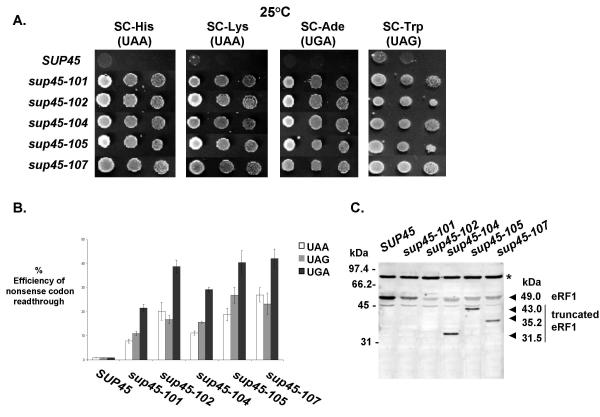
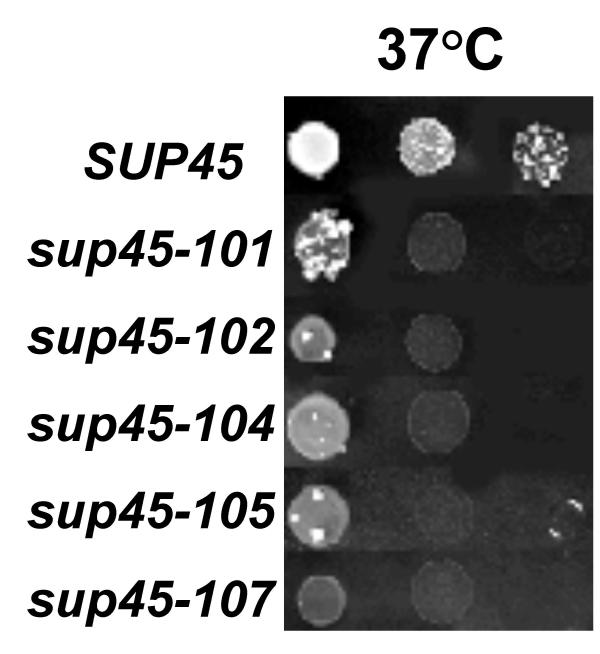
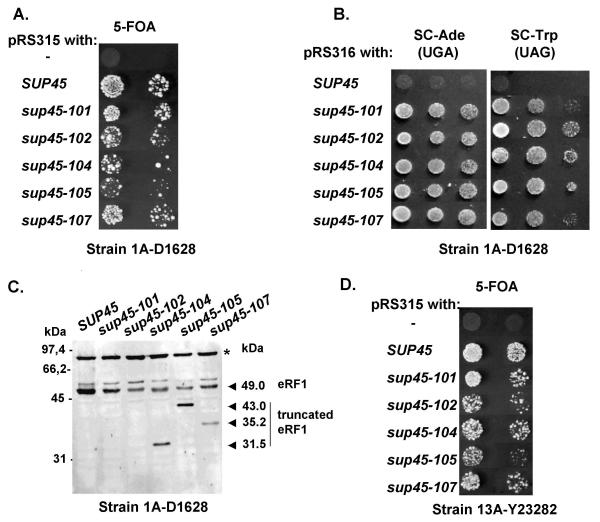
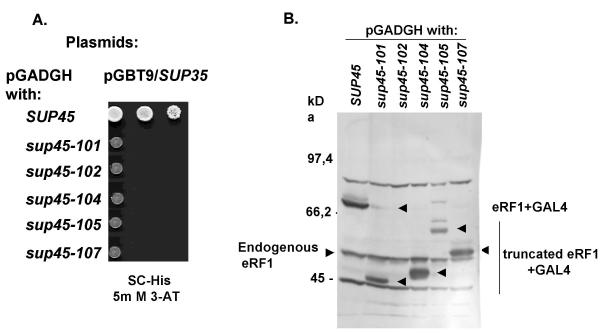
Results: We have isolated five sup45-n (n from nonsense) mutations that cause nonsense substitutions in the following amino acid positions of eRF1: Y53 --> UAA, E266 --> UAA, L283 --> UAA, L317 --> UGA, E385 --> UAA. We found that full-length eRF1 protein is present in all mutants, although in decreased amounts. All mutations are situated in a weak termination context. All these sup45-n mutations are viable in different genetic backgrounds, however their viability increases after growth in the absence of wild-type allele. Any of sup45-n mutations result in temperature sensitivity (37 degrees C). Most of the sup45-n mutations lead to decreased spore viability and spores bearing sup45-n mutations are characterized by limited budding after germination leading to formation of microcolonies of 4-20 cells.
Conclusions: Nonsense mutations in the essential gene SUP45 can be isolated in the absence of tRNA nonsense suppressors.
Figures
Similar articles
-
[Viable nonsense mutants for the SUP45 gene in the yeast Saccharomyces cerevisiae are lethal at increased temperature].Genetika. 2007 Oct;43(10):1363-71. Genetika. 2007. PMID: 18069340 Russian.
-
Regulation of release factor expression using a translational negative feedback loop: a systems analysis.RNA. 2012 Dec;18(12):2320-34. doi: 10.1261/rna.035113.112. Epub 2012 Oct 25. RNA. 2012. PMID: 23104998 Free PMC article.
-
Gene Amplification as a Mechanism of Yeast Adaptation to Nonsense Mutations in Release Factor Genes.Genes (Basel). 2021 Dec 19;12(12):2019. doi: 10.3390/genes12122019. Genes (Basel). 2021. PMID: 34946968 Free PMC article.
-
Modulation of efficiency of translation termination in Saccharomyces cerevisiae.Prion. 2014;8(3):247-60. doi: 10.4161/pri.29851. Epub 2014 Nov 1. Prion. 2014. PMID: 25486049 Free PMC article. Review.
-
Eukaryotic release factors (eRFs) history.Biol Cell. 2003 May-Jun;95(3-4):195-209. doi: 10.1016/s0248-4900(03)00035-2. Biol Cell. 2003. PMID: 12867083 Review.
Cited by
-
Genome Sequencing and Comparative Analysis of Saccharomyces cerevisiae Strains of the Peterhof Genetic Collection.PLoS One. 2016 May 6;11(5):e0154722. doi: 10.1371/journal.pone.0154722. eCollection 2016. PLoS One. 2016. PMID: 27152522 Free PMC article.
-
Microbial Community Structure in Ancient European Arctic Peatlands.Plants (Basel). 2022 Oct 13;11(20):2704. doi: 10.3390/plants11202704. Plants (Basel). 2022. PMID: 36297728 Free PMC article.
-
Nonsense Mutations in the Yeast SUP35 Gene Affect the [PSI+] Prion Propagation.Int J Mol Sci. 2020 Feb 28;21(5):1648. doi: 10.3390/ijms21051648. Int J Mol Sci. 2020. PMID: 32121268 Free PMC article.
-
Chromosome-level genome assembly and structural variant analysis of two laboratory yeast strains from the Peterhof Genetic Collection lineage.G3 (Bethesda). 2021 Apr 15;11(4):jkab029. doi: 10.1093/g3journal/jkab029. G3 (Bethesda). 2021. PMID: 33677552 Free PMC article.
-
Distinct paths to stop codon reassignment by the variant-code organisms Tetrahymena and Euplotes.Mol Cell Biol. 2006 Jan;26(2):438-47. doi: 10.1128/MCB.26.2.438-447.2006. Mol Cell Biol. 2006. PMID: 16382136 Free PMC article.
References
-
- Bertram G, Innes S, Minella O, Richardson J, Stansfield I. Endless possibilities: translation termination and stop codon recognition. Microbiology. 2001;147:255–269. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
