

Selective activation of the c-Jun NH2-terminal protein kinase signaling pathway by stimulatory KIR in the absence of KARAP/DAP12 in CD4+ T cells
- PMID: 12591902
- PMCID: PMC2193867
- DOI: 10.1084/jem.20020383
Selective activation of the c-Jun NH2-terminal protein kinase signaling pathway by stimulatory KIR in the absence of KARAP/DAP12 in CD4+ T cells
Abstract
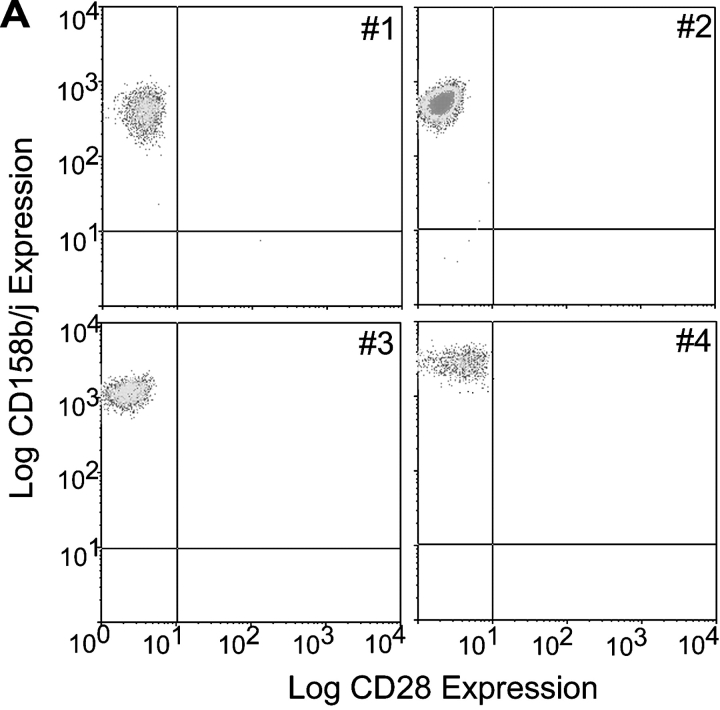
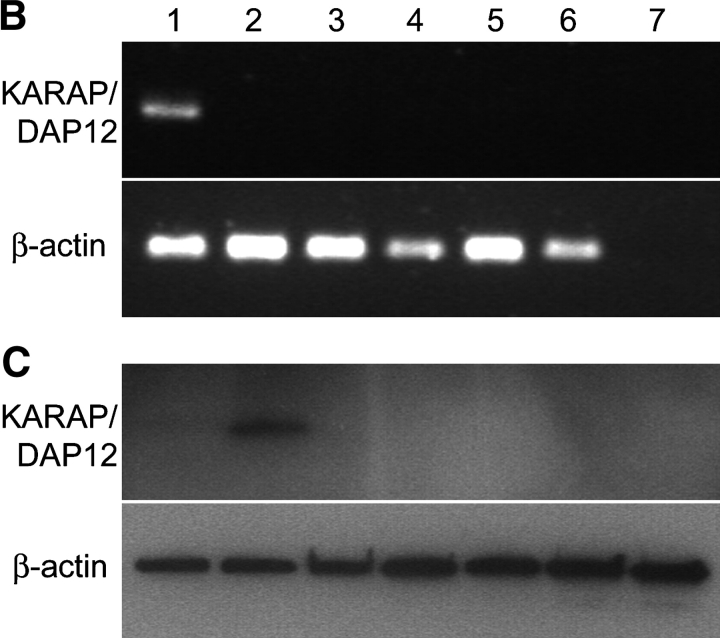
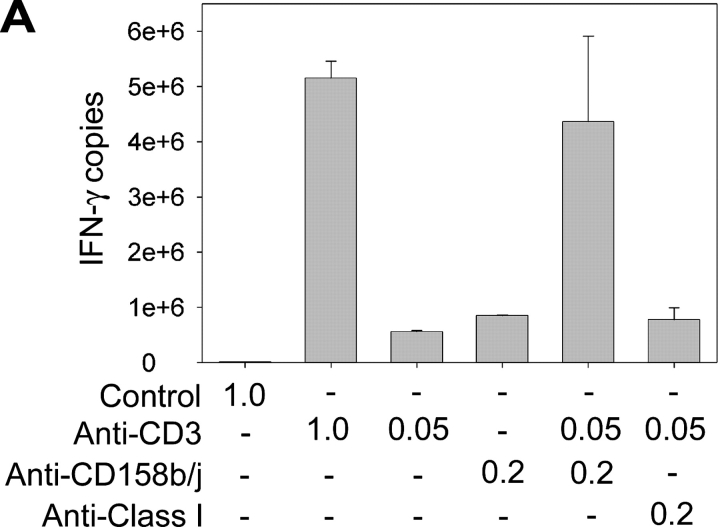
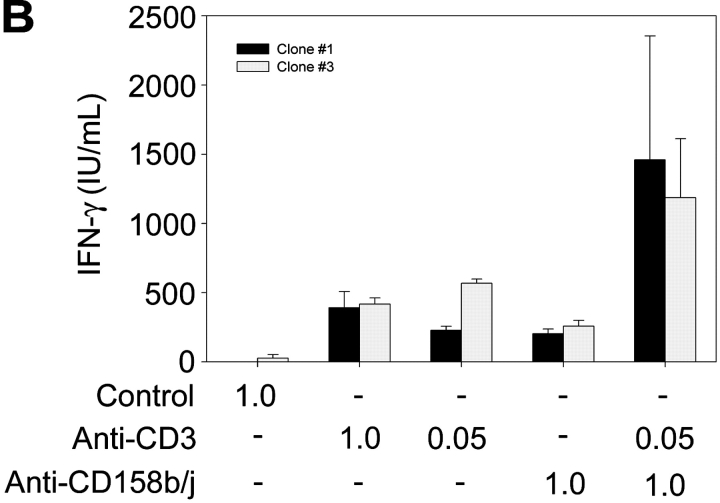
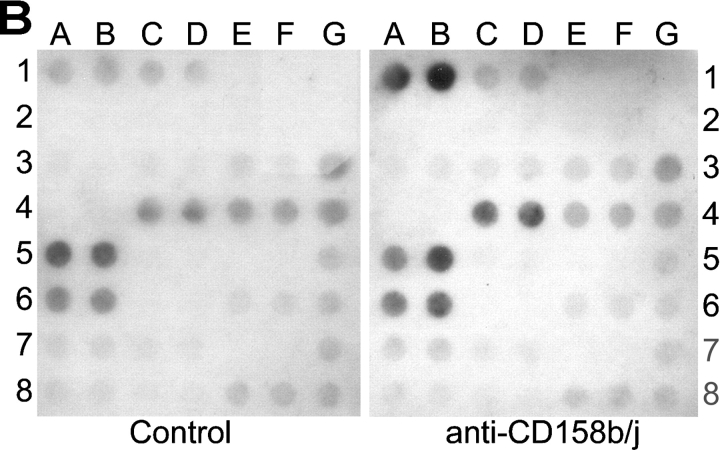
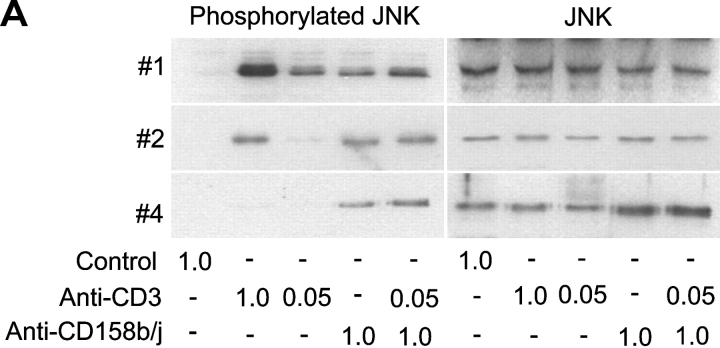
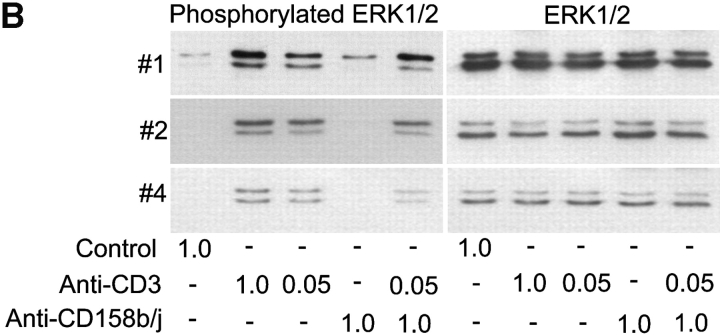
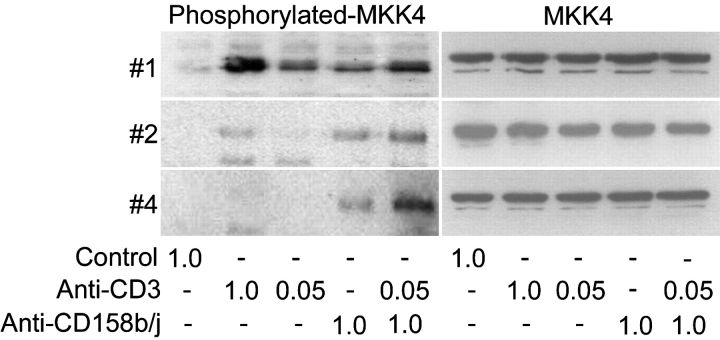
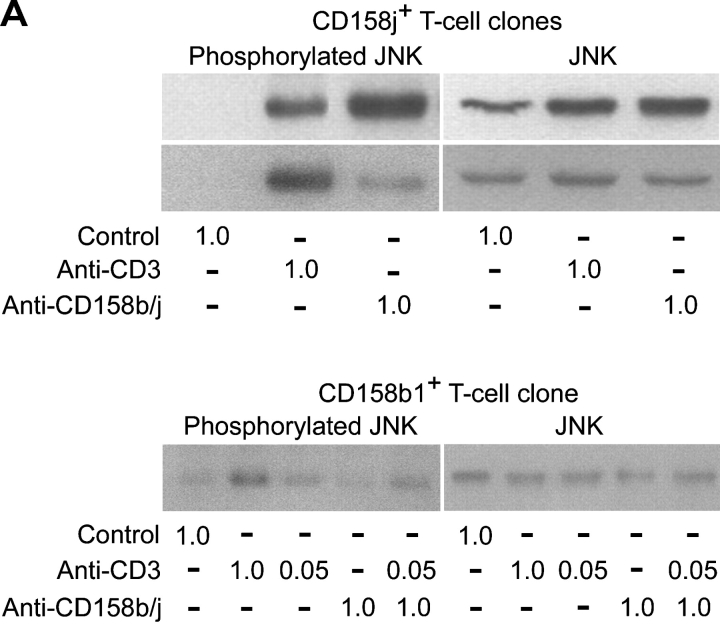
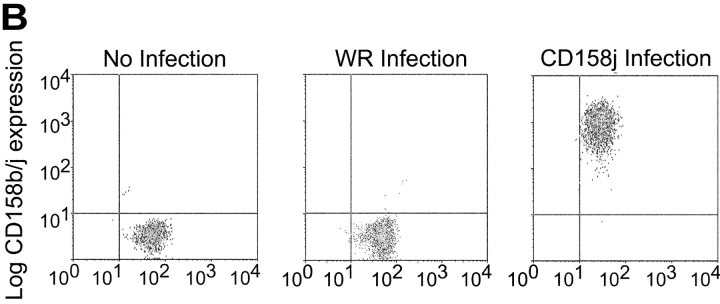
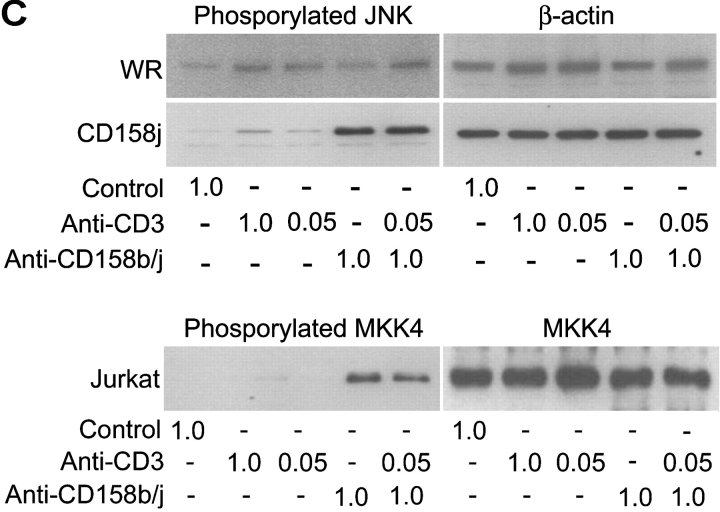
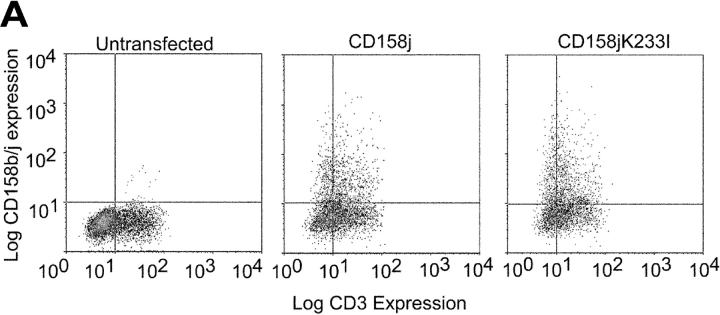
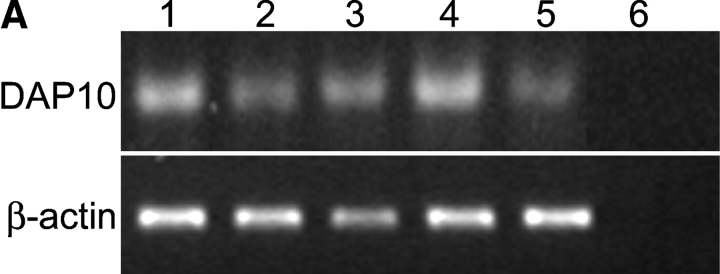
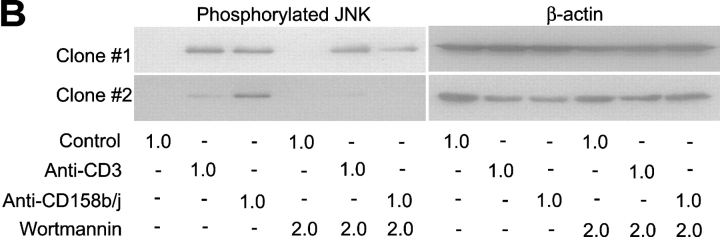
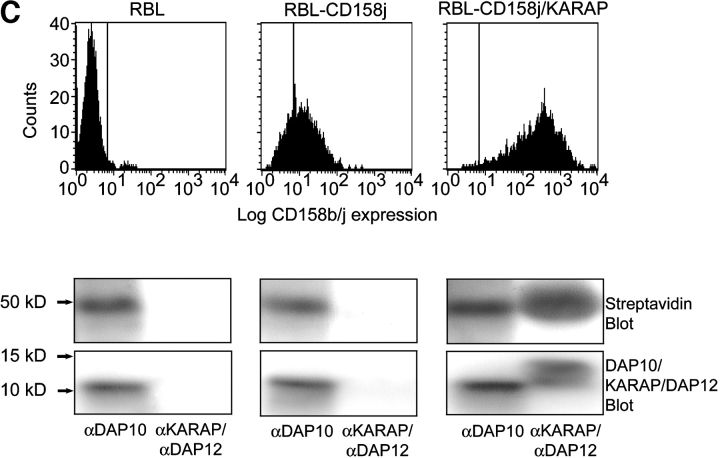
Activation of CD4(+) T cells is governed by interplay between stimulatory and inhibitory receptors; predominance of stimulatory signals favors autoimmune reactions. In patients with rheumatoid arthritis, expression of the critical costimulatory molecule, CD28, is frequently lost. Instead, CD4(+)CD28(null) T cells express killer immunoglobulin-like receptors (KIRs) with a preferential expression of the stimulatory receptor, CD158j. The frequency of CD4(+)CD28(null) T cells in rheumatoid arthritis (RA) correlates with the risk for more severe disease. Moreover, the KIR2DS2 gene, which encodes for CD158j, is a genetic risk factor for rheumatoid vasculitis. CD158j signals through the adaptor molecule, KARAP/DAP12, to positively regulate cytotoxic activity in NK cells. However, the majority of CD4(+)CD28(null) T cell clones lacked the expression of KARAP/DAP12. Despite the absence of KARAP/DAP12, CD158j was functional and augmented interferon-gamma production after T cell receptor stimulation. Cross-linking of CD158j resulted in selective phosphorylation of c-Jun NH(2)-terminal protein kinase (JNK) and its upstream kinase, MKK4 that led to the expression of ATF-2 and c-Jun, all in the absence of extracellular signal-regulated kinase (ERK)1/2 phosphorylation. Mutation of the lysine residue within the transmembrane domain of CD158j abolished JNK activation, suggesting that an alternate adaptor molecule was being used. CD4(+)CD28(null) T cells expressed DAP10 and inhibition of phosphatidylinositol 3-kinase, which acts downstream of DAP10, inhibited JNK activation; however, no interaction of DAP10 with CD158j could be detected. Our data suggest that CD158j in T cells functions as a costimulatory molecule through the JNK pathway independent of KARAP/DAP12 and DAP10. Costimulation by CD158j may contribute to the autoreactivity of CD4(+)CD28(null) T cells in RA.
Figures




References
-
- Olcese, L., P. Lang, F. Vely, A. Cambiaggi, D. Marguet, M. Blery, K.L. Hippen, R. Biassoni, A. Moretta, L. Moretta, et al. 1996. Human and mouse killer-cell inhibitory receptors recruit PTP1C and PTP1D protein tyrosine phosphatases. J. Immunol. 156:4531–4534. - PubMed
-
- Binstadt, B.A., K.M. Brumbaugh, C.J. Dick, A.M. Scharenberg, B.L. Williams, M. Colonna, L.L. Lanier, J.P. Kinet, R.T. Abraham, and P.J. Leibson. 1996. Sequential involvement of Lck and SHP-1 with MHC-recognizing receptors on NK cells inhibits FcR-initiated tyrosine kinase activation. Immunity. 5:629–638. - PubMed
-
- Ljunggren, H.G., and K. Karre. 1990. In search of the ‘missing self’: MHC molecules and NK cell recognition. Immunol. Today. 11:237–244. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
