

Identification and characterization of a Streptococcus pyogenes operon involved in binding of hemoproteins and acquisition of iron
- PMID: 12595414
- PMCID: PMC148835
- DOI: 10.1128/IAI.71.3.1042-1055.2003
Identification and characterization of a Streptococcus pyogenes operon involved in binding of hemoproteins and acquisition of iron
Abstract
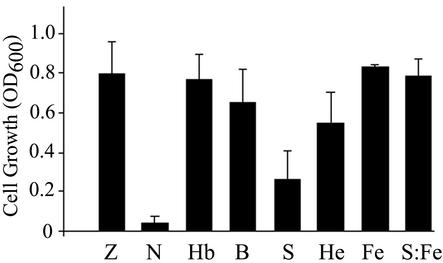
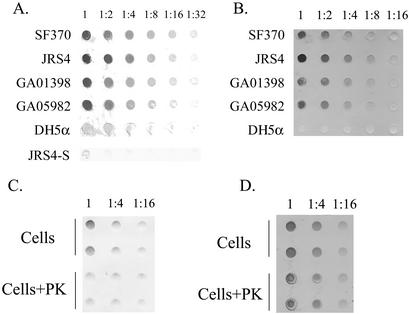
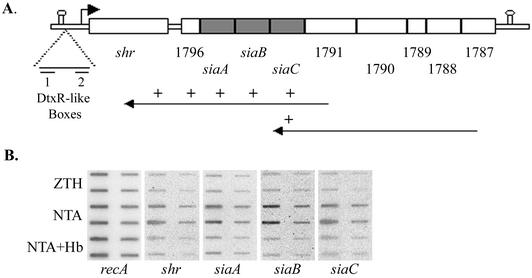
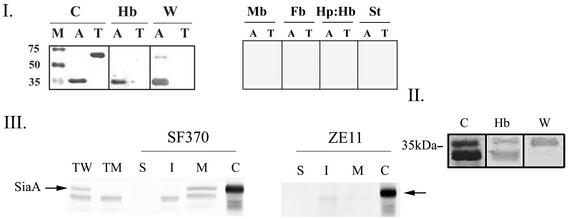
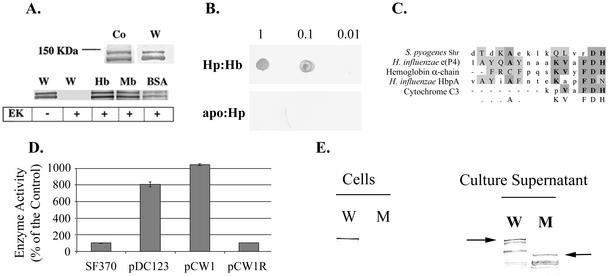
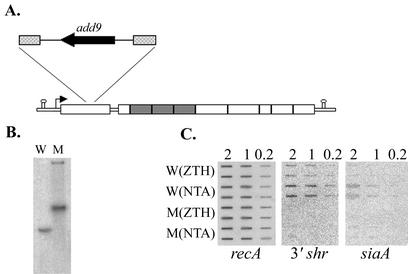
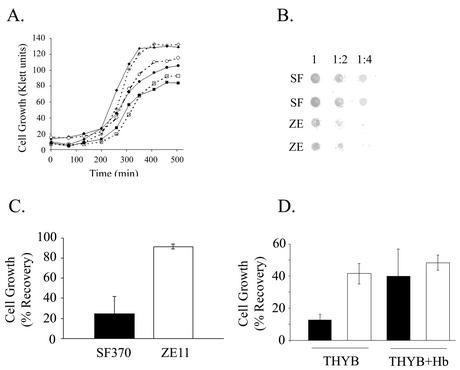
The hemolytic Streptococcus pyogenes can use a variety of heme compounds as an iron source. In this study, we investigate hemoprotein utilization by S. pyogenes. We demonstrate that surface proteins contribute to the binding of hemoproteins to S. pyogenes. We identify an ABC transporter from the iron complex family named sia for streptococcal iron acquisition, which consists of a lipoprotein (siaA), membrane permease (siaB), and ATPase (siaC). The sia transporter is part of a highly conserved, iron regulated, 10-gene operon. SiaA, which was localized to the cell membrane, could specifically bind hemoglobin. The operon's first gene encodes a novel bacterial protein that bound hemoglobin, myoglobin, heme-albumin, and hemoglobin-haptoglobin (but not apo-haptoglobin) and therefore was named Shr, for streptococcal hemoprotein receptor. PhoZ fusion and Western blot analysis showed that Shr has a leader peptide and is found in both membrane-bound and soluble forms. An M1 SF370 strain with a polar mutation in shr was more resistant to streptonigrin and hydrogen peroxide, suggesting decreased iron uptake. The addition of hemoglobin to the culture medium increased cell resistance to hydrogen peroxide in SF370 but not in the mutant, implying the sia operon may be involved in hemoglobin-dependent resistance to oxidative stress. The shr mutant demonstrated reduced hemoglobin binding, though cell growth in iron-depleted medium supplemented with hemoglobin, whole blood, or ferric citrate was not affected, suggesting additional systems are involved in hemoglobin utilization. SiaA and Shr are the first hemoprotein receptors identified in S. pyogenes; their possible role in iron capture is discussed.
Figures
References
-
- Al-Tawfiq, J. A., K. R. Fortney, B. P. Katz, A. F. Hood, C. Elkins, and S. M. Spinola. 2000. An isogenic hemoglobin receptor-deficient mutant of Haemophilus ducreyi is attenuated in the human model of experimental infection. J. Infect. Dis. 181:1049-1054. - PubMed
-
- Altman, P. L., and D. D. Katz. 1961. Blood and other body fluids. Federation of American Societies for Experimental Biology, Washington, D.C.
-
- Brown, J. S., S. M. Gilliland, and D. W. Holden. 2001. A Streptococcus pneumoniae pathogenicity island encoding an ABC transporter involved in iron uptake and virulence. Mol. Microbiol. 40:572-585. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
