

Studies of NMDA receptor function and stoichiometry with truncated and tandem subunits
- PMID: 12598603
- PMCID: PMC6742241
- DOI: 10.1523/JNEUROSCI.23-04-01151.2003
Studies of NMDA receptor function and stoichiometry with truncated and tandem subunits
Abstract
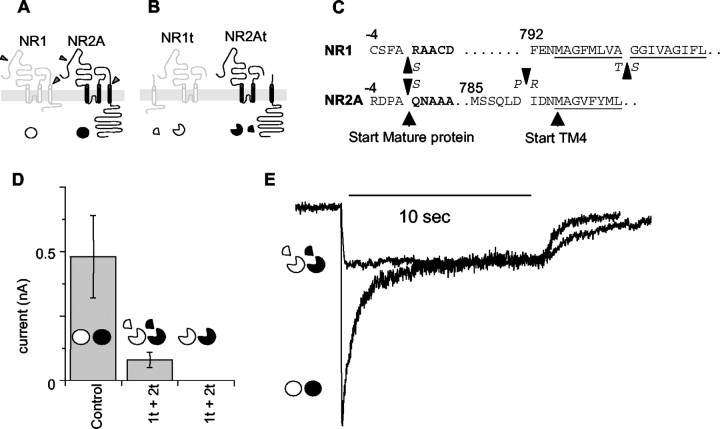
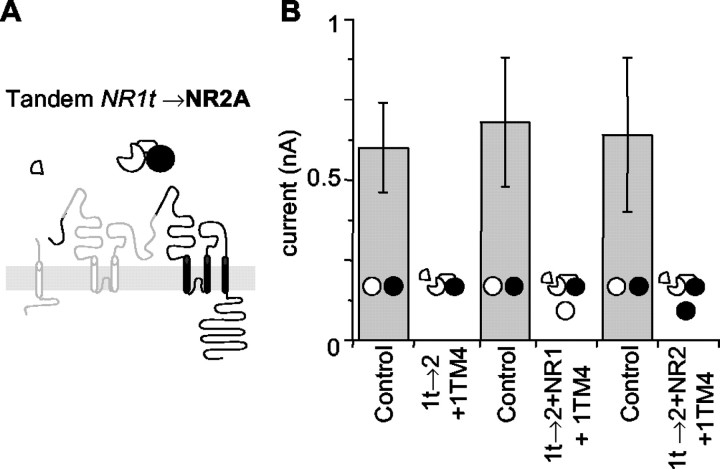
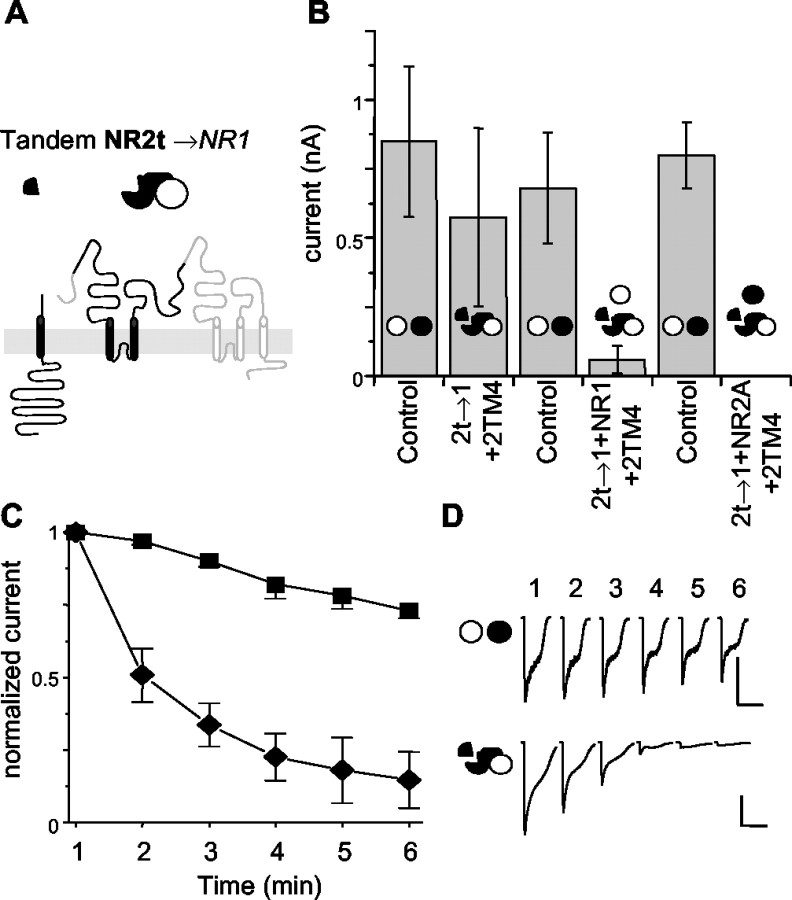
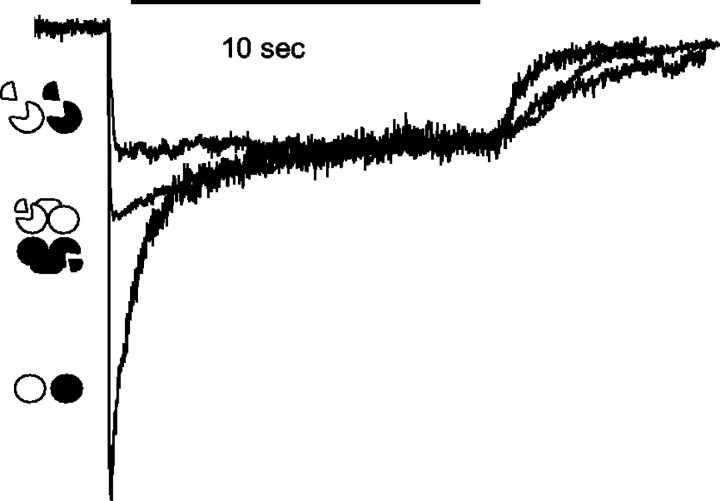
The subunits that compose eukaryotic glutamate ion channel receptors have three transmembrane domains (TMs) and terminate with intracellular tails that are important for controlling channel expression and localization. Truncation of NMDA receptor subunits before the final TM showed that this TM and intracellular tail region are necessary to form functional channels. However, it is shown here that these truncated subunits may be partially rescued by coexpressing the final TM and tail as a separate protein. The whole-cell currents so produced are somewhat lower than with full-length subunits, and they do not show the sag characteristic of currents from channels containing NR1 and NR2A subunits in the continued presence of an agonist. In addition, these truncated subunits were joined to full-length subunits to generate tandems. The functional expression of these tandems confirmed the tetrameric structure of NMDA receptors and also suggested that the subunits making up NMDA receptors are arranged as a dimer of dimers in the receptors with a 1-1-2-2 orientation of the subunits in the channel, and not in an alternating pattern of subunits around the pore. These results may redirect future studies into the mechanism of binding and gating in these receptors toward schemes including dimers, and may also be relevant to studies of glutamate receptor ion channels in general.
Figures


References
-
- Armstrong N, Gouaux E. Mechanisms for activation and antagonism of an AMPA-sensitive glutamate receptor: crystal structures of the GluR2 ligand binding core. Neuron. 2000;28:165–181. - PubMed
-
- Baumann SW, Baur R, Sigel E. Subunit arrangement of γ-aminobutyric acid type A receptors. J Biol Chem. 2001;276:36275–36280. - PubMed
-
- Behe P, Stern P, Wyllie DJ, Nassar M, Schoepfer R, Colquhoun D. Determination of NMDA NR1 subunit copy number in recombinant NMDA receptors. Proc R Soc Lond B Biol Sci. 1995;262:205–213. - PubMed
-
- Bigge CF. Ionotropic glutamate receptors. Curr Opin Chem Biol. 1999;3:441–447. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials