

Functional role of C-terminal cytoplasmic tail of rat vanilloid receptor 1
- PMID: 12598622
- PMCID: PMC6742269
- DOI: 10.1523/JNEUROSCI.23-04-01340.2003
Functional role of C-terminal cytoplasmic tail of rat vanilloid receptor 1
Abstract
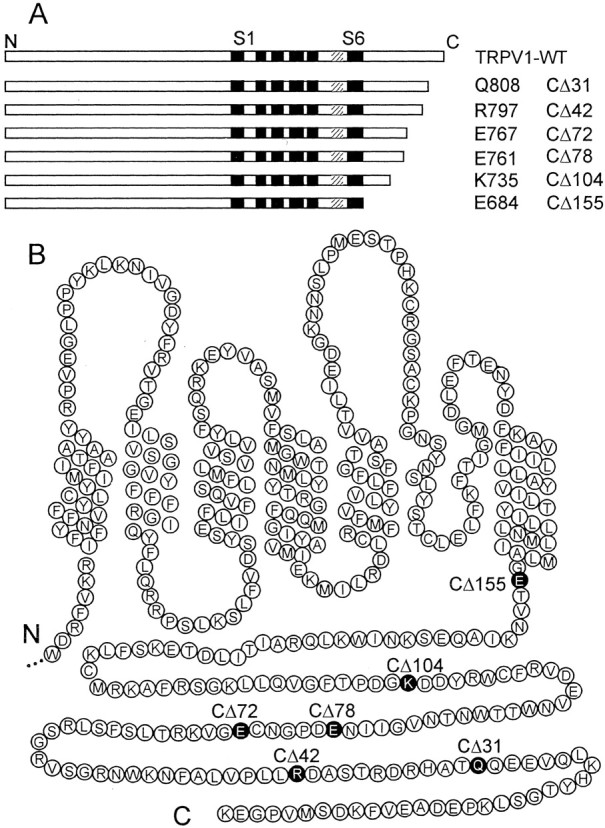
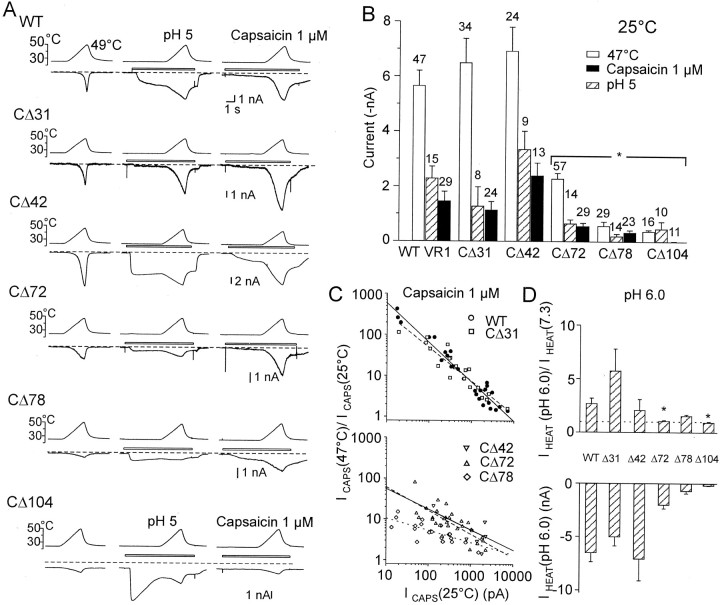
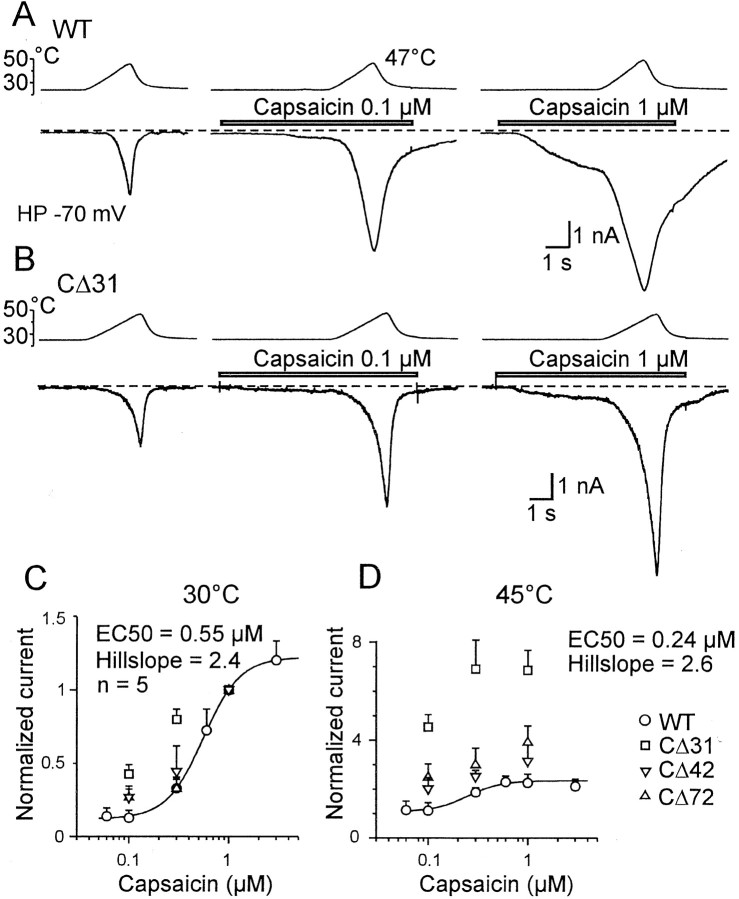
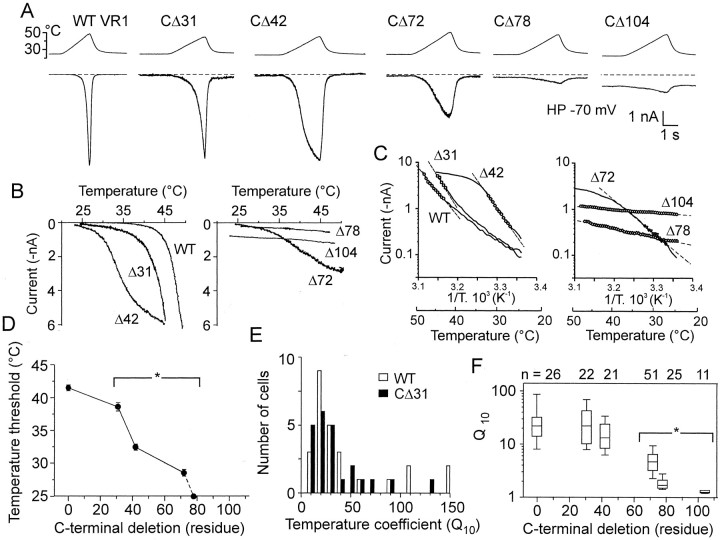
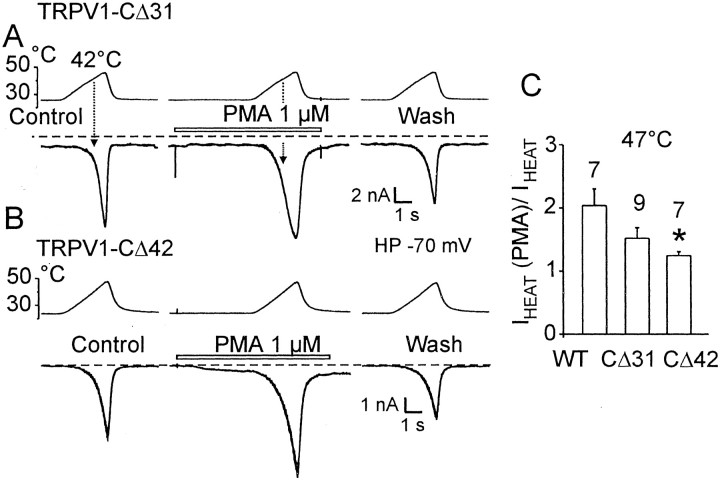
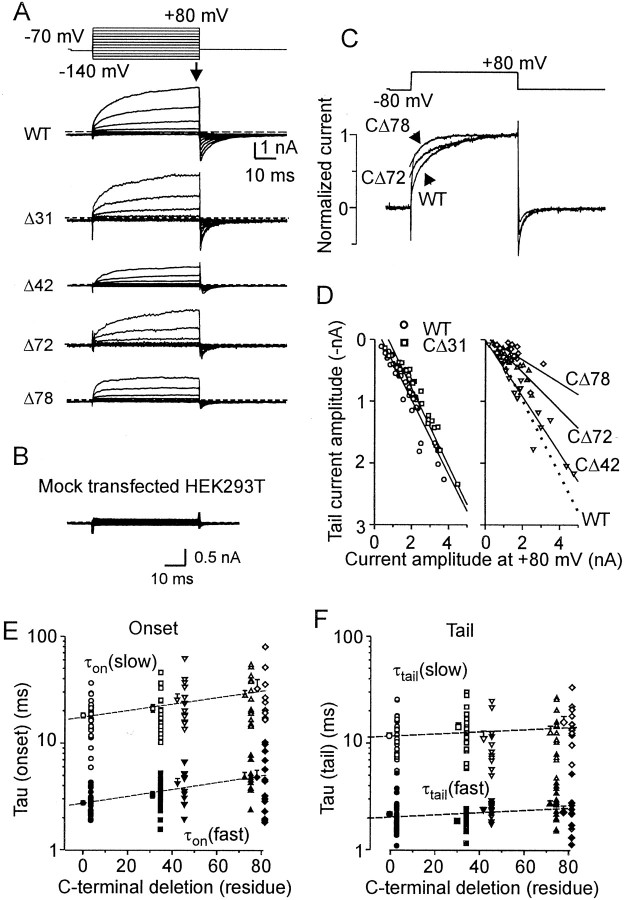
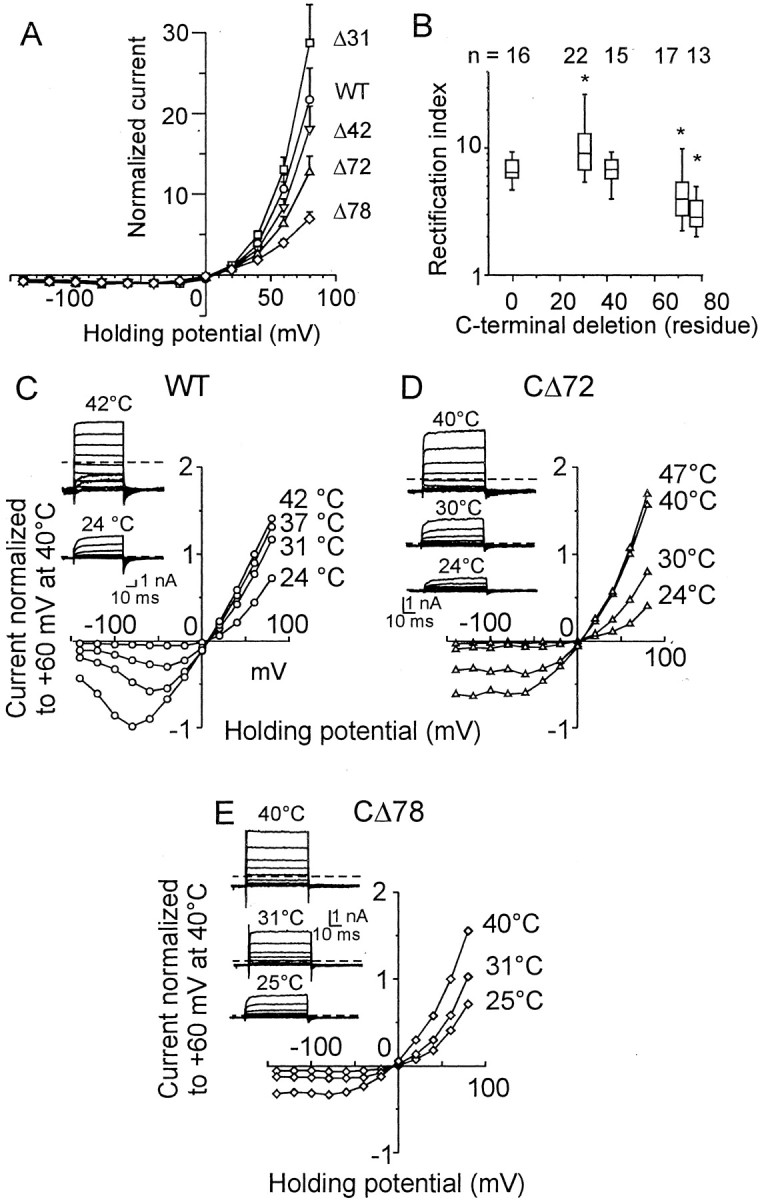
The vanilloid receptor [transient receptor potential (TRP)V1, also known as VR1] is a member of the TRP channel family. These receptors share a significant sequence homology, a similar predicted structure with six transmembrane-spanning domains (S1-S6), a pore-forming region between S5 and S6, and the cytoplasmically oriented C- and N-terminal regions. Although structural/functional studies have identified some of the key amino acids influencing the gating of the TRPV1 ion channel, the possible contributions of terminal regions to vanilloid receptor function remain elusive. In the present study, C-terminal truncations of rat TRPV1 have been constructed to characterize the contribution of the cytoplasmic C-terminal region to TRPV1 function and to delineate the minimum amount of C tail necessary to form a functional channel. The truncation of 31 residues was sufficient to induce changes in functional properties of TRPV1 channel. More pronounced effects of C-terminal truncation were seen in mutants lacking the final 72 aa. These changes were characterized by a decline of capsaicin-, pH-, and heat-sensitivity; progressive reduction of the activation thermal threshold (from 41.5 to 28.6 degrees C); and slowing of the activation rate of heat-evoked membrane currents (Q10 from 25.6 to 4.7). The voltage-induced currents of the truncated mutants exhibited a slower onset, markedly reduced outward rectification, and significantly smaller peak tail current amplitudes. Truncation of the entire TRPV1 C-terminal domain (155 residues) resulted in a nonfunctional channel. These results indicate that the cytoplasmic COOH-terminal domain strongly influences the TRPV1 channel activity, and that the distal half of this structural domain confers specific thermal sensitivity.
Figures
References
-
- Bentley GN, Brooks MA, O'Neill CA, Findlay JB. Determinants of potassium channel assembly localised within the cytoplasmic C-terminal domain of Kv2.1. Biochim Biophys Acta. 1999;1418:176–184. - PubMed
-
- Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D. The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature. 1997;389:816–824. - PubMed
-
- Caterina MJ, Rosen TA, Tominaga M, Brake AJ, Julius D. A capsaicin-receptor homologue with a high threshold for noxious heat. Nature. 1999;398:436–441. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases