

Rho kinase inhibition enhances axonal regeneration in the injured CNS
- PMID: 12598630
- PMCID: PMC6742251
- DOI: 10.1523/JNEUROSCI.23-04-01416.2003
Rho kinase inhibition enhances axonal regeneration in the injured CNS
Abstract
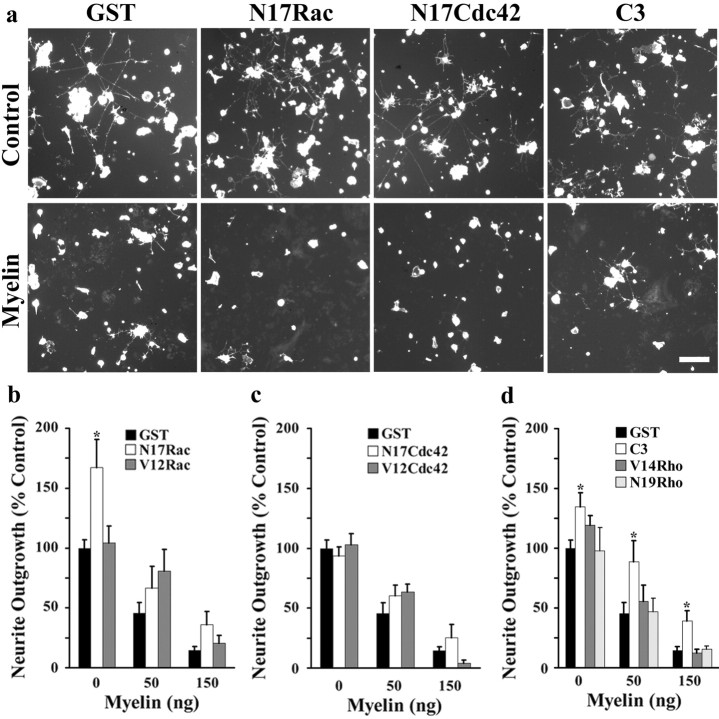
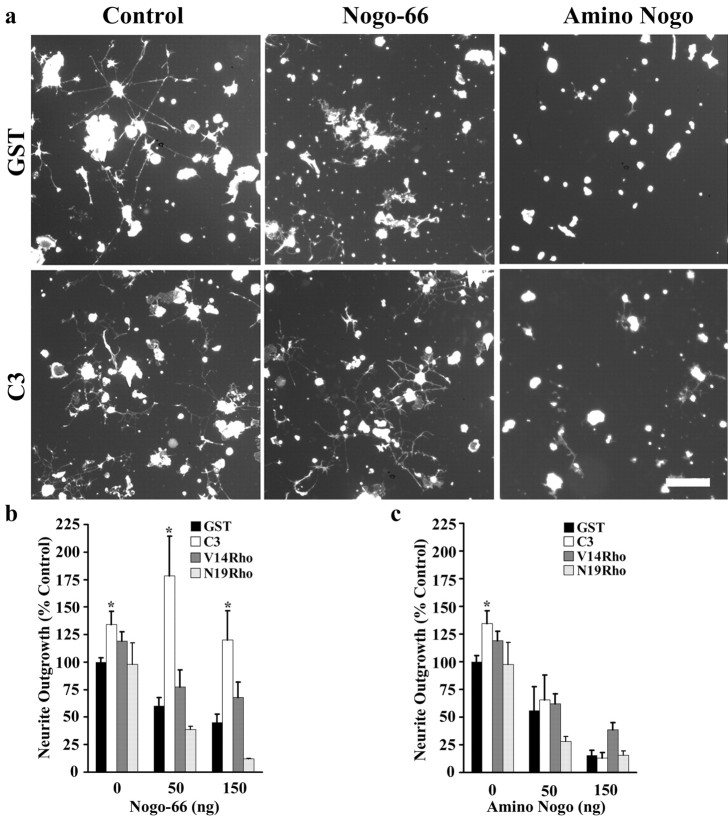
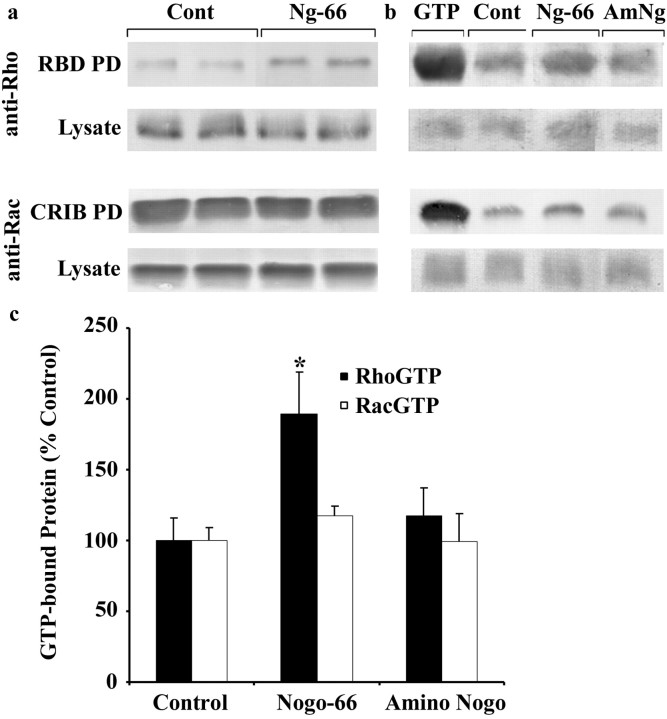
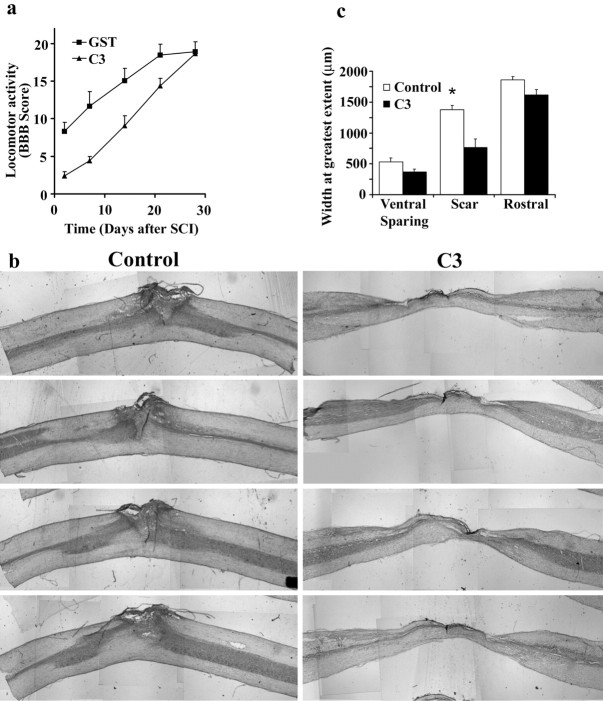
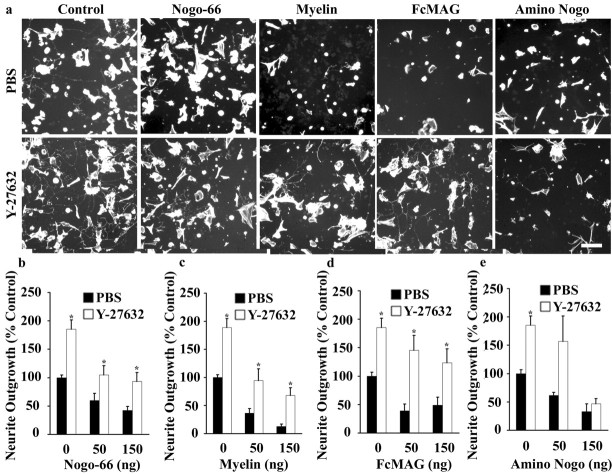
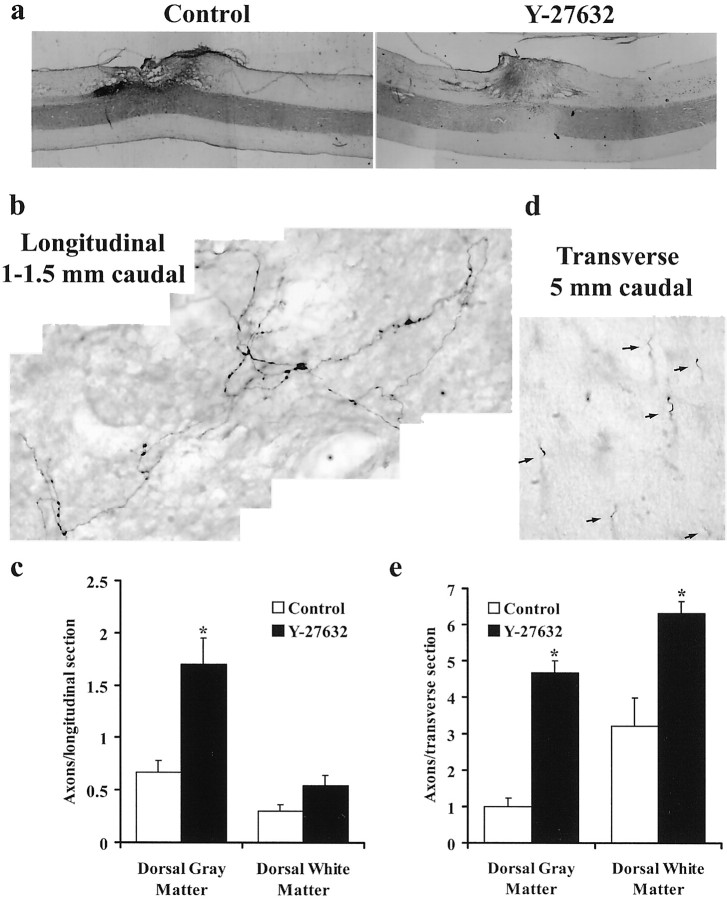
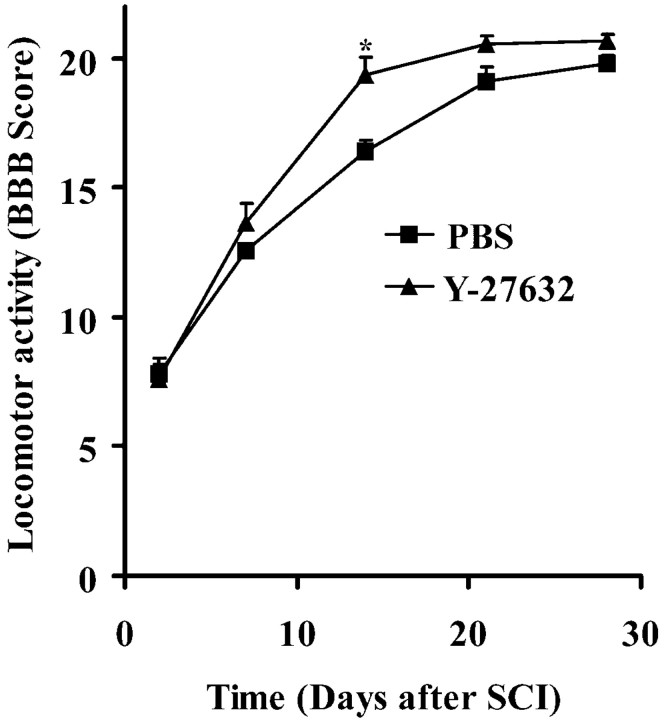
Myelin-associated inhibitors limit axonal regeneration in the injured brain and spinal cord. A common target of many neurite outgrowth inhibitors is the Rho family of small GTPases. Activation of Rho and a downstream effector of Rho, p160ROCK, inhibits neurite outgrowth. Here, we demonstrate that Rho is directly activated by the myelin-associated inhibitor Nogo-66. Using a binding assay to measure Rho activity, we detected increased levels of GTP Rho in PC12 and dorsal root ganglion (DRG) cell lysates after Nogo-66 stimulation. Rho activity levels were not affected by Amino-Nogo stimulation. Rho inactivation with C3 transferase promotes neurite outgrowth of chick DRG neurons in vitro, but with the delivery method used here, it fails to promote neurite outgrowth after corticospinal tract (CST) lesions in the adult rat. Inhibition of p160ROCK with Y-27632 also promotes neurite outgrowth on myelin-associated inhibitors in vitro. Furthermore, Y-27632 enhances sprouting of CST fibers in vivo and accelerates locomotor recovery after CST lesions in adult rats.
Figures
References
-
- Amano M, Mukai H, Ono Y, Chihara K, Matsui T, Hamajima Y, Okawa K, Iwamatsu A, Kaibuchi K. Identification of a putative target for Rho as the serine-threonine kinase protein kinase N. Science. 1996;271:648–650. - PubMed
-
- Amano M, Chihara K, Kimura K, Fukata Y, Nakamura N, Matsuura Y, Kaibuchi K. Formation of actin stress fibers and focal adhesions enhanced by Rho-kinase. Science. 1997;275:1308–1311. - PubMed
-
- Aspenstrom P, Lindberg U, Hall A. Two GTPases, Cdc42 and Rac, bind directly to a protein implicated in the immunodeficiency disorder Wiskott–Aldrich syndrome. Curr Biol. 1996;6:70–75. - PubMed
-
- Bagrodia S, Taylor SJ, Creasy CL, Chernoff J, Cerione RA. Identification of a mouse p21Cdc42/Rac activated kinase. J Biol Chem. 1995;270:22731–22737. - PubMed
-
- Basso DM, Beattie MS, Bresnahan JC. A sensitive and reliable locomotor rating scale for open field testing in rats. J Neurotrauma. 1995;12:1–21. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous