

The ATP hydrolyzing transcription activator phage shock protein F of Escherichia coli: identifying a surface that binds sigma 54
- PMID: 12601152
- PMCID: PMC151331
- DOI: 10.1073/pnas.0537525100
The ATP hydrolyzing transcription activator phage shock protein F of Escherichia coli: identifying a surface that binds sigma 54
Abstract
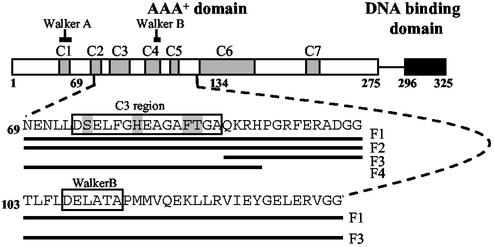
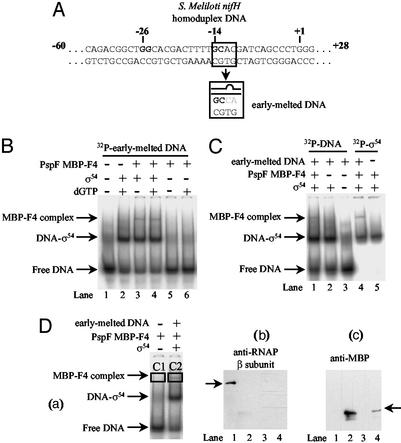
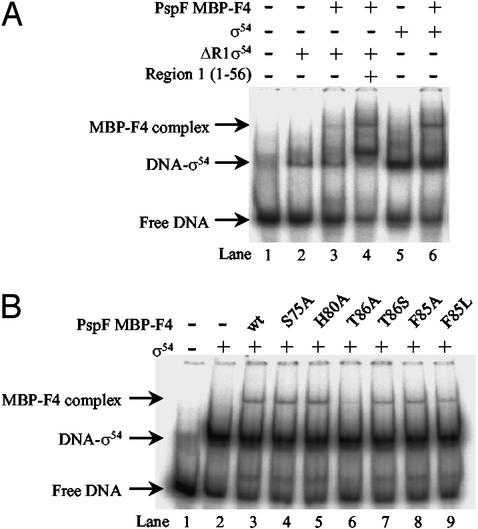
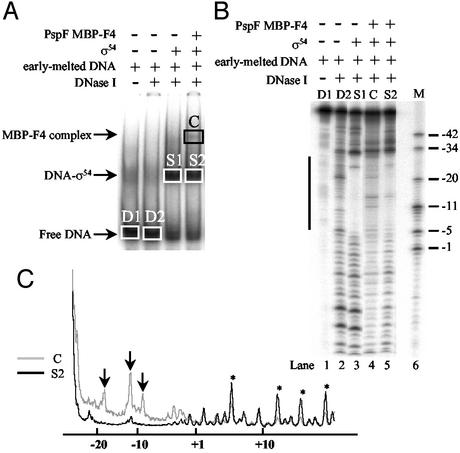
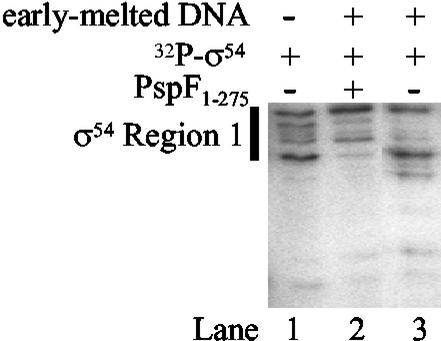
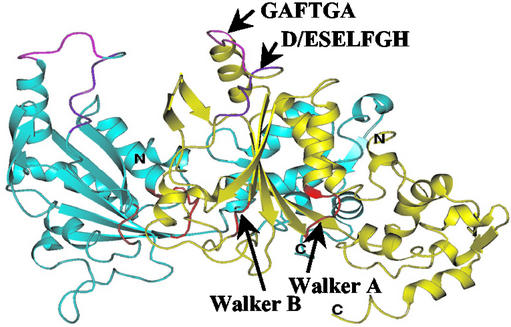
Members of the protein family called ATPases associated with various cellular activities (AAA(+)) play a crucial role in transforming chemical energy into biological events. AAA(+) proteins are complex molecular machines and typically form ring-shaped oligomeric complexes that are crucial for ATPase activity and mechanism of action. The Escherichia coli transcription activator phage shock protein F (PspF) is an AAA(+) mechanochemical enzyme that functions to sense and relay the energy derived from nucleoside triphosphate hydrolysis to catalyze transcription by the sigma(54)-RNA polymerase. Closed promoter complexes formed by the sigma(54)-RNA polymerase are substrates for the action of PspF. By using a protein fragmentation approach, we identify here at least one sigma(54)-binding surface in the PspF AAA(+) domain. Results suggest that ATP hydrolysis by PspF is coupled to the exposure of at least one sigma(54)-binding surface. This nucleotide hydrolysis-dependent presentation of a substrate binding surface can explain why complexes that form between sigma(54) and PspF are transient and could be part of a mechanism used generally by other AAA(+) proteins to regulate activity.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
