

Transgenerational inheritance of epigenetic states at the murine Axin(Fu) allele occurs after maternal and paternal transmission
- PMID: 12601169
- PMCID: PMC151376
- DOI: 10.1073/pnas.0436776100
Transgenerational inheritance of epigenetic states at the murine Axin(Fu) allele occurs after maternal and paternal transmission
Abstract
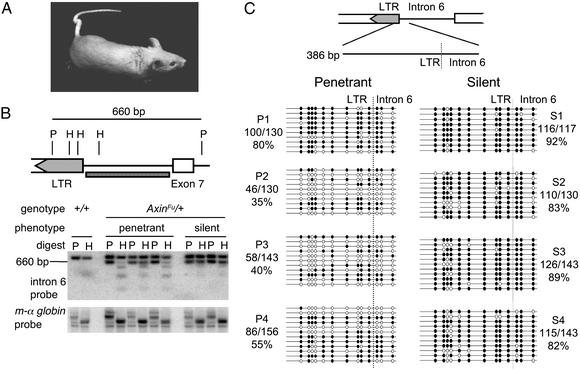
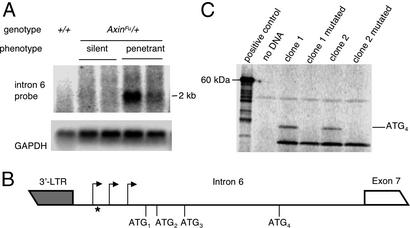
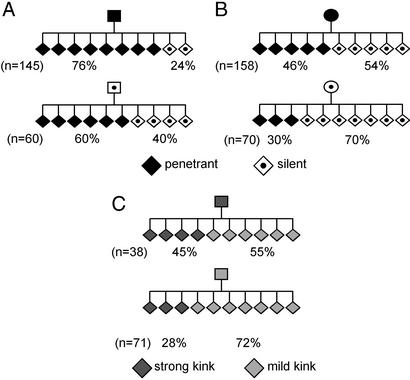
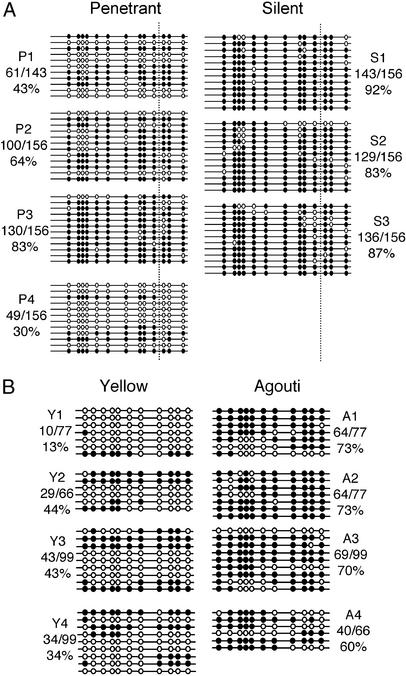
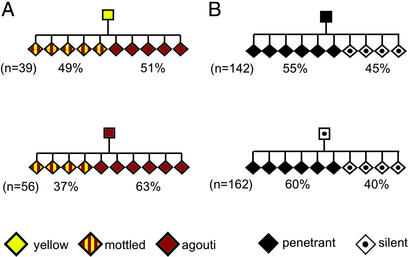
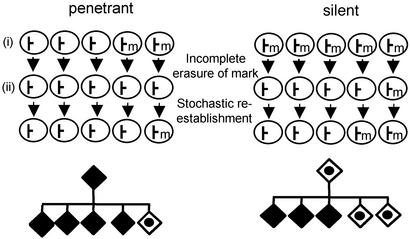
Phenotypic variation that cannot be explained by genetic or environmental heterogeneity has intrigued geneticists for decades. The molecular basis of this phenomenon, however, is largely a mystery. Axin-fused (Axin(Fu)), first identified in 1937, is a classic example of a mammalian allele displaying extremely variable expression states. Here we demonstrate that the presence or absence of its characteristic phenotype, a kinked tail, correlates with differential DNA methylation at a retrotransposon within Axin(Fu) and identify mutant transcripts arising adjacent to the retrotransposon LTR that are likely to be causative of the phenotype. Furthermore, the epigenetic state at Axin(Fu) can be inherited transgenerationally after both maternal and paternal transmission. This is in contrast to epigenetic inheritance at the murine agouti-viable yellow (A(vy)) allele, which occurs through the female only. Unlike the egg, the sperm contributes very little (if any) cytoplasm to the zygote, and therefore paternal inheritance at Axin(Fu) argues against the possibility that the effects are due to cytoplasmic or metabolic influences. Consistent with the idea of transgenerational inheritance of epigenetic marks, we find that the methylation state of Axin(Fu) in mature sperm reflects the methylation state of the allele in the somatic tissue of the animal, suggesting that it does not undergo epigenetic reprogramming during gametogenesis. Finally, we show that epigenetic inheritance is influenced by strain background. These findings enable us to propose a model for transgenerational epigenetic inheritance in mammals.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
