

Active-site residues of Escherichia coli DNA gyrase required in coupling ATP hydrolysis to DNA supercoiling and amino acid substitutions leading to novobiocin resistance
- PMID: 12604539
- PMCID: PMC149296
- DOI: 10.1128/AAC.47.3.1037-1046.2003
Active-site residues of Escherichia coli DNA gyrase required in coupling ATP hydrolysis to DNA supercoiling and amino acid substitutions leading to novobiocin resistance
Abstract
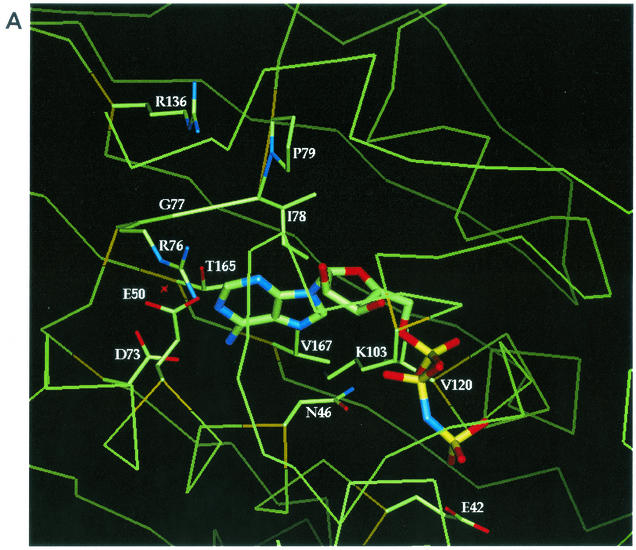
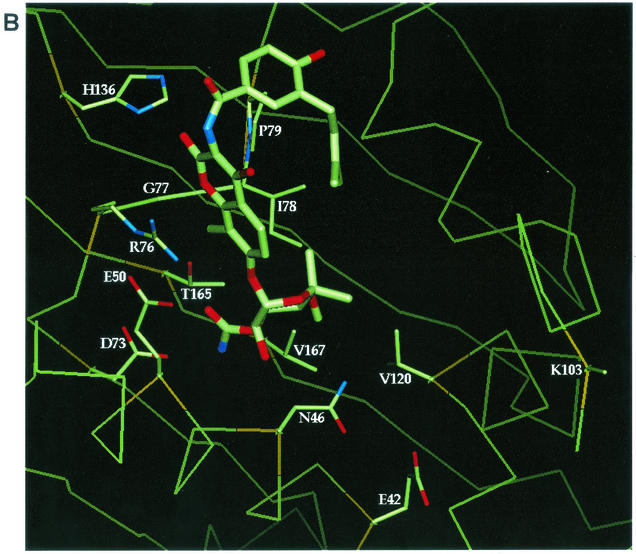
DNA gyrase is a bacterial type II topoisomerase which couples the free energy of ATP hydrolysis to the introduction of negative supercoils into DNA. Amino acids in proximity to bound nonhydrolyzable ATP analog (AMP. PNP) or novobiocin in the gyrase B (GyrB) subunit crystal structures were examined for their roles in enzyme function and novobiocin resistance by site-directed mutagenesis. Purified Escherichia coli GyrB mutant proteins were complexed with the gyrase A subunit to form the functional A(2)B(2) gyrase enzyme. Mutant proteins with alanine substitutions at residues E42, N46, E50, D73, R76, G77, and I78 had reduced or no detectable ATPase activity, indicating a role for these residues in ATP hydrolysis. Interestingly, GyrB proteins with P79A and K103A substitutions retained significant levels of ATPase activity yet demonstrated no DNA supercoiling activity, even with 40-fold more enzyme than the wild-type enzyme, suggesting that these amino acid side chains have a role in the coupling of the two activities. All enzymes relaxed supercoiled DNA to the same extent as the wild-type enzyme did, implying that only ATP-dependent reactions were affected. Mutant genes were examined in vivo for their abilities to complement a temperature-sensitive E. coli gyrB mutant, and the activities correlated well with the in vitro activities. We show that the known R136 novobiocin resistance mutations bestow a significant loss of inhibitor potency in the ATPase assay. Four new residues (D73, G77, I78, and T165) that, when changed to the appropriate amino acid, result in both significant levels of novobiocin resistance and maintain in vivo function were identified in E. coli.
Figures
Similar articles
-
Crystal structures of Escherichia coli topoisomerase IV ParE subunit (24 and 43 kilodaltons): a single residue dictates differences in novobiocin potency against topoisomerase IV and DNA gyrase.Antimicrob Agents Chemother. 2004 May;48(5):1856-64. doi: 10.1128/AAC.48.5.1856-1864.2004. Antimicrob Agents Chemother. 2004. PMID: 15105144 Free PMC article.
-
Energy coupling in DNA gyrase and the mechanism of action of novobiocin.Proc Natl Acad Sci U S A. 1978 Oct;75(10):4838-42. doi: 10.1073/pnas.75.10.4838. Proc Natl Acad Sci U S A. 1978. PMID: 368801 Free PMC article.
-
Alteration of Escherichia coli topoisomerase IV to novobiocin resistance.Antimicrob Agents Chemother. 2003 Mar;47(3):941-7. doi: 10.1128/AAC.47.3.941-947.2003. Antimicrob Agents Chemother. 2003. PMID: 12604525 Free PMC article.
-
The interaction between coumarin drugs and DNA gyrase.Mol Microbiol. 1993 Aug;9(4):681-6. doi: 10.1111/j.1365-2958.1993.tb01728.x. Mol Microbiol. 1993. PMID: 8231802 Review.
-
DNA gyrase and the supercoiling of DNA.Science. 1980 Feb 29;207(4434):953-60. doi: 10.1126/science.6243420. Science. 1980. PMID: 6243420 Review.
Cited by
-
3D-QSAR and docking studies of flavonoids as potent Escherichia coli inhibitors.Sci Rep. 2016 Apr 6;6:23634. doi: 10.1038/srep23634. Sci Rep. 2016. PMID: 27049530 Free PMC article.
-
Functional interactions between gyrase subunits are optimized in a species-specific manner.J Biol Chem. 2020 Feb 21;295(8):2299-2312. doi: 10.1074/jbc.RA119.010245. Epub 2020 Jan 17. J Biol Chem. 2020. PMID: 31953321 Free PMC article.
-
Real-time visualization of photochemically induced fluorescence of 8-halogenated quinolones: lomefloxacin, clinafloxacin and Bay3118 in live human HaCaT keratinocytes.Photochem Photobiol. 2010 Jul-Aug;86(4):792-7. doi: 10.1111/j.1751-1097.2010.00741.x. Epub 2010 May 13. Photochem Photobiol. 2010. PMID: 20492567 Free PMC article.
-
Exploiting bacterial DNA gyrase as a drug target: current state and perspectives.Appl Microbiol Biotechnol. 2011 Nov;92(3):479-97. doi: 10.1007/s00253-011-3557-z. Epub 2011 Sep 9. Appl Microbiol Biotechnol. 2011. PMID: 21904817 Free PMC article. Review.
-
Ligand-Based Virtual Screening for Discovery of Indole Derivatives as Potent DNA Gyrase ATPase Inhibitors Active against Mycobacterium tuberculosis and Hit Validation by Biological Assays.J Chem Inf Model. 2024 Aug 12;64(15):5991-6002. doi: 10.1021/acs.jcim.4c00511. Epub 2024 Jul 12. J Chem Inf Model. 2024. PMID: 38993154 Free PMC article.
References
-
- Akasaka, T., M. Tanaka, A. Yamaguchi, and K. Sato. 2001. Type II topoisomerase mutation in fluoroquinolone-resistant clinical strains of Pseudomonas aeruginosa isolated in 1998 and 1999: role of target enzyme in mechanism of fluoroquinolone resistance. Antimicrob. Agents Chemother. 45:2263-2268. - PMC - PubMed
-
- Ali, J. A., A. P. Jackson, A. J. Howells, and A. Maxwell. 1993. The 43-kilodalton N-terminal fragment of the DNA gyrase B protein hydrolyzes ATP and binds coumarin drugs. Biochemistry 32:2717-2724. - PubMed
-
- Brino, L., A. Urzhumtsev, M. Mousli, C. Bronner, A. Mitschler, P. Oudet, and D. Moras. 2000. Dimerization of Escherichia coli DNA-gyrase B provides a structural mechanism for activating the ATPase catalytic center. J. Biol. Chem. 275:9468-9475. - PubMed
-
- Contreras, A., and A. Maxwell. 1992. GyrB mutations which confer coumarin resistance also affect DNA supercoiling and ATP hydrolysis by Escherichia coli DNA gyrase. Mol. Microbiol. 6:1617-1624. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
