

Intrabodies based on intracellular capture frameworks that bind the RAS protein with high affinity and impair oncogenic transformation
- PMID: 12606568
- PMCID: PMC150339
- DOI: 10.1093/emboj/cdg106
Intrabodies based on intracellular capture frameworks that bind the RAS protein with high affinity and impair oncogenic transformation
Abstract
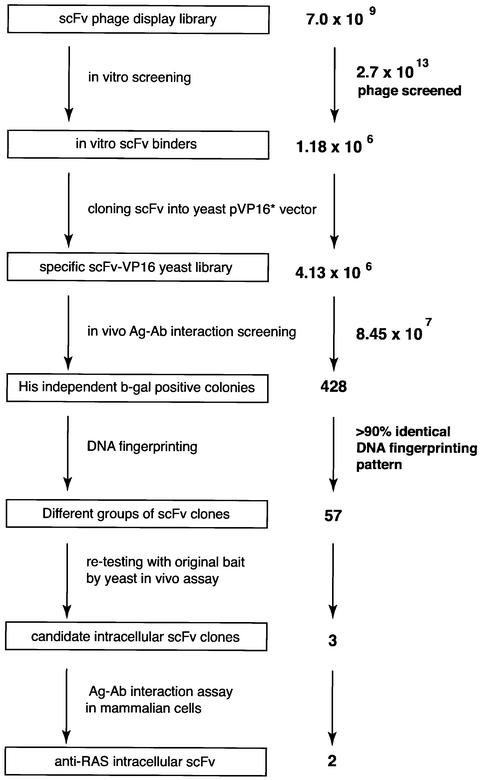
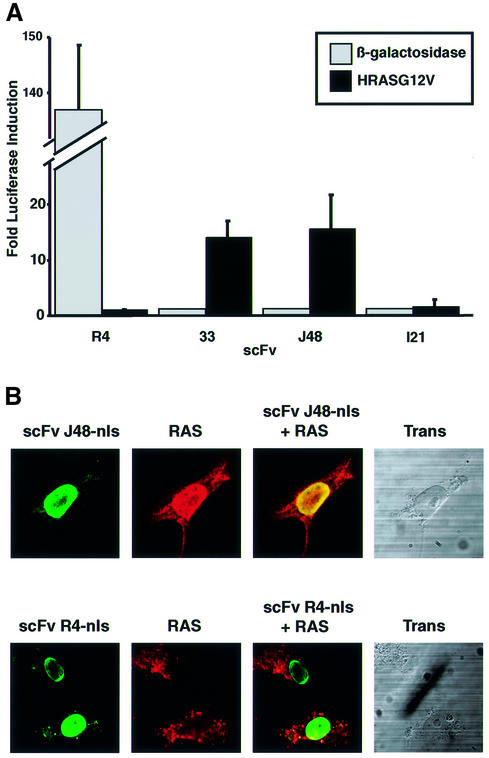
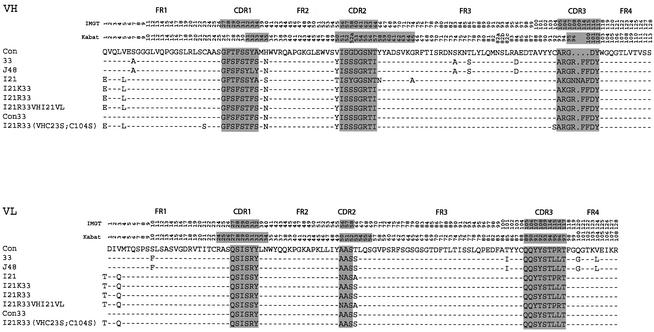
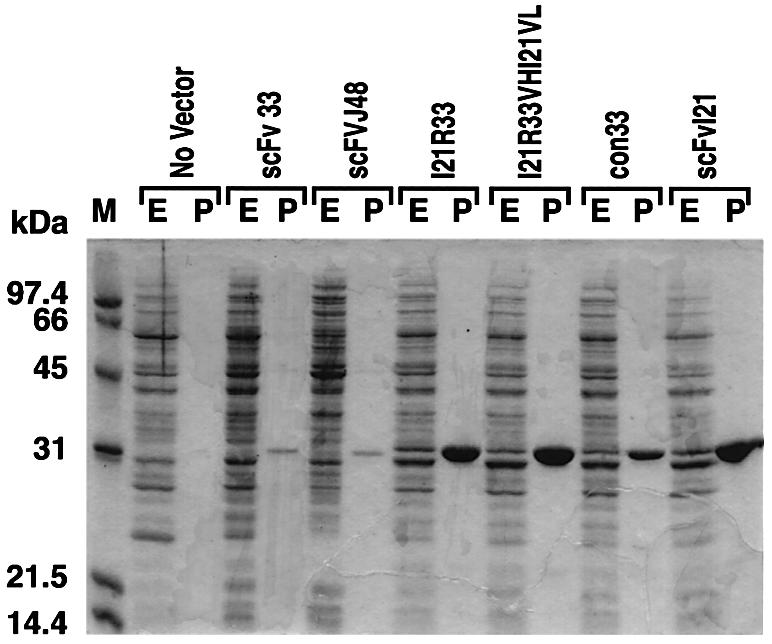
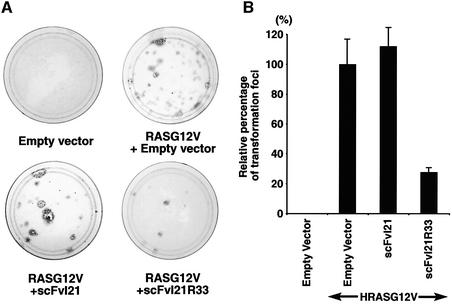
We have applied in vivo intracellular antibody capture (IAC) technology to isolate human intrabodies which bind to the oncogenic RAS protein. IAC facilitates the capture of antibody fragments, in this case single-chain Fvs (scFvs), which tolerate reducing environments, such as the cytoplasm of cancer cells. Three anti-RAS scFvs with different affinity, solubility and intracellular binding activity were characterized. The anti-RAS scFvs with highest affinity were expressed relatively poorly in mammalian cells, and greater soluble expression was achieved by mutating the antibody framework to canonical consensus scaffolds, previously derived from IAC, without losing antigen specificity. Mutagenesis experiments showed that the consensus scaffolds are functional as intrabody fragments without an intra-domain disulfide bond. Furthermore, we could convert an intrabody which does not bind RAS in mammalian cells into a high-affinity reagent capable of inhibiting RAS-mediated NIH 3T3 transformation by exchanging VH and VL complementarity-determining regions onto its consensus scaffold. These data show that the consensus scaffold is a robust framework by which to improve intrabody function.
Figures
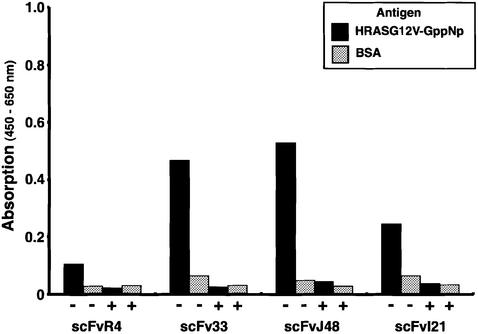
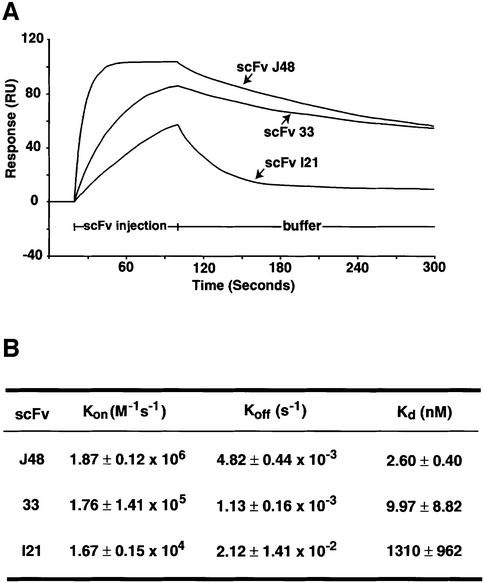
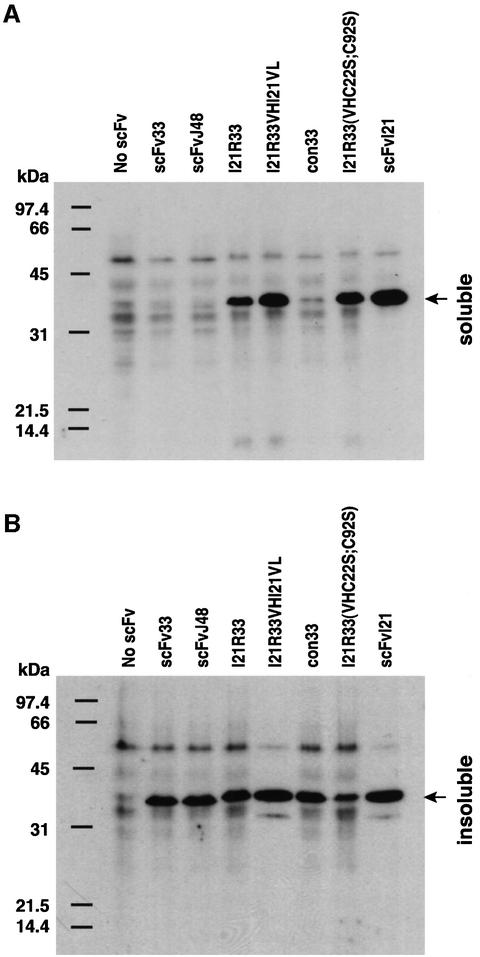
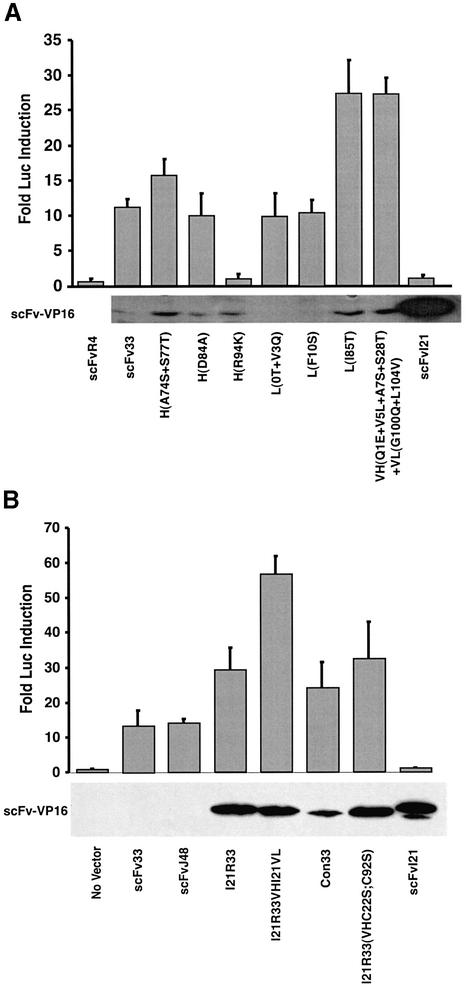
References
-
- Adjei A.A. (2001) Blocking oncogenic Ras signaling for cancer therapy. J. Natl Cancer Inst., 93, 1062–1074. - PubMed
-
- Biocca S., Pierandrei-Amaldi,P. and Cattaneo,A. (1993) Intracellular expression of anti-p21ras single chain Fv fragments inhibits meiotic maturation of Xenopus oocytes. Biochem. Biophys. Res. Commun., 197, 422–427. - PubMed
-
- Biocca S., Pierandrei-Amaldi,P., Campioni,N. and Cattaneo,A. (1994) Intracellular immunization with cytosolic recombinant antibodies. Biotechnology, 12, 396–399. - PubMed
-
- Biocca S., Ruberti,F., Tafani,M., Pierandrei-Amaldi,P. and Cattaneo,A. (1995) Redox state of single chain Fv fragments targeted to the endoplasmic reticulum, cytosol and mitochondria. Biotechnology, 13, 1110–1115. - PubMed
-
- Bird R.E. et al. (1988) Single-chain antigen-binding proteins. Science, 242, 423–426. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
