

The human chromokinesin Kid is a plus end-directed microtubule-based motor
- PMID: 12606572
- PMCID: PMC150335
- DOI: 10.1093/emboj/cdg102
The human chromokinesin Kid is a plus end-directed microtubule-based motor
Abstract
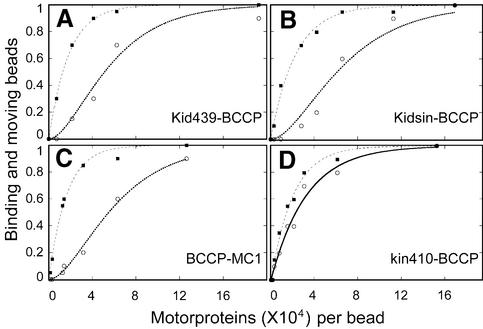
Kid is a kinesin-like DNA-binding protein known to be involved in chromosome movement during mitosis, although its actual motor function has not been demonstrated. Here, we describe the initial characterization of Kid as a microtubule-based motor using optical trapping microscopy. A bacterially expressed fusion protein consisting of a truncated Kid fragment (amino acids 1-388 or 1-439) is indeed an active microtubule motor with an average speed of approximately 160 nm/s, and the polarity of movement is plus end directed. We could not detect processive movement of either monomeric Kid or dimerizing chimeric Kid; however, low levels of processivity (a few steps) cannot be detected with our method. These results are consistent with Kid having a role in chromosome congression in vivo, where it would be responsible for the polar ejection forces acting on the chromosome arms.
Figures
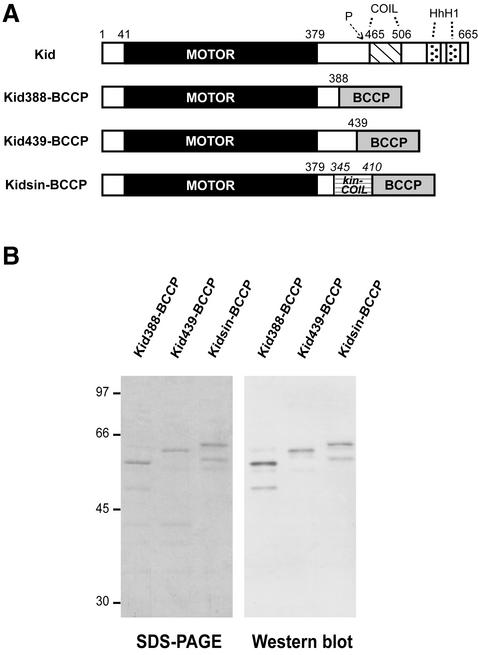
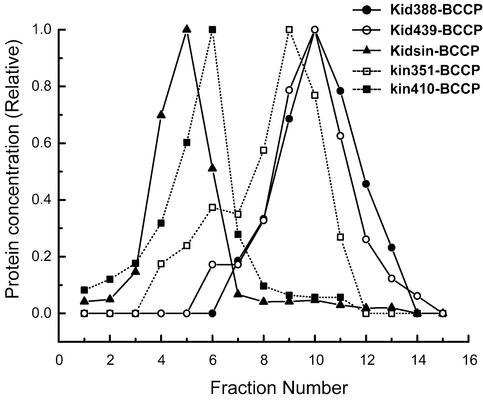
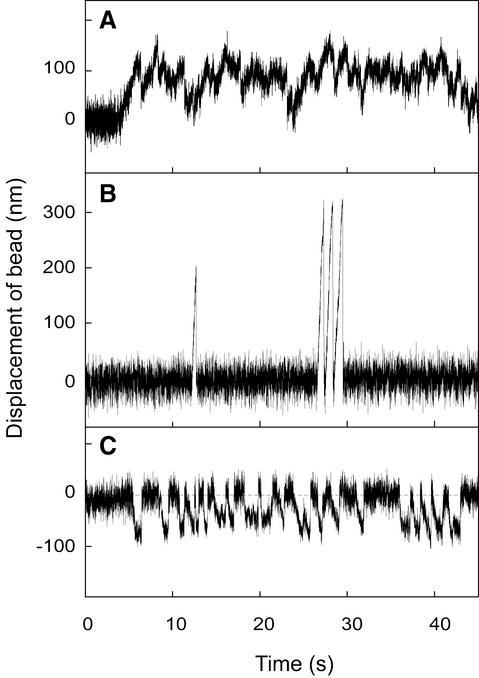
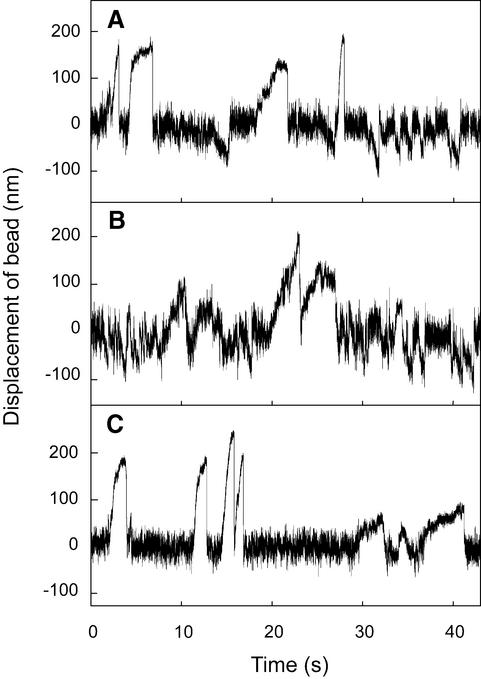
References
-
- Afshar K., Barton,N.R., Hawley,R.S. and Goldstein,L.S. (1995b) DNA binding and meiotic chromosomal localization of the Drosophila nod kinesin-like protein. Cell, 81, 129–138. - PubMed
-
- Antonio C., Ferby,I., Wilhelm,H., Jones,M., Karsenti,E., Nebreda,A.R. and Vernos,I. (2000) Xkid, a chromokinesin required for chromosome alignment on the metaphase plate. Cell, 102, 425–435. - PubMed
-
- Berliner E., Mahtani,H.K., Karki,S., Chu,L.F., Cronan,J.E. and Gelles,J.,Jr (1994) Microtubule movement by a biotinated kinesin bound to streptavidin-coated surface. J. Biol. Chem., 269, 8610–8615. - PubMed
-
- Chandra R., Salmon,E.D., Erickson,H.P., Lockhart,A. and Endow,S.A. (1993) Structural and functional domains of the Drosophila ncd microtubule motor protein. J. Biol. Chem., 268, 9005–9013. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
