

Lineage-restricted retention of a primitive immunoglobulin heavy chain isotype within the Dipnoi reveals an evolutionary paradox
- PMID: 12606718
- PMCID: PMC151370
- DOI: 10.1073/pnas.0538029100
Lineage-restricted retention of a primitive immunoglobulin heavy chain isotype within the Dipnoi reveals an evolutionary paradox
Abstract
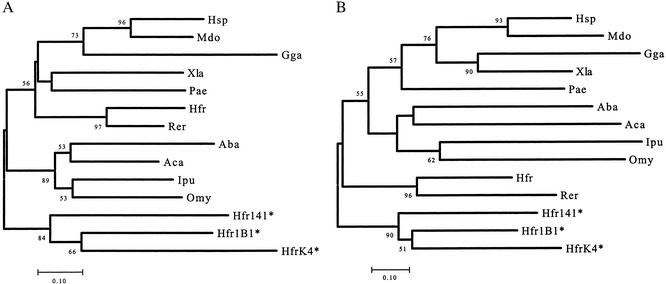
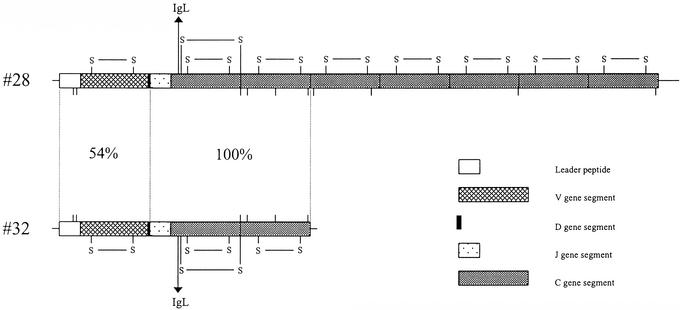
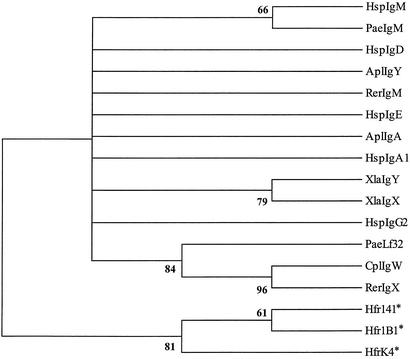
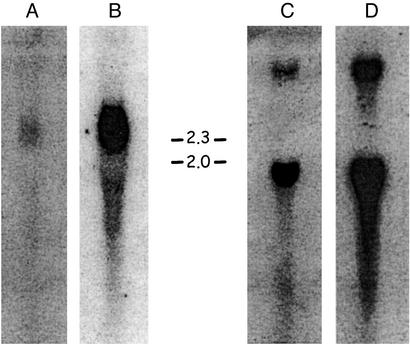
The lineage leading to lungfishes is one of the few major jawed vertebrate groups in which Ig heavy chain isotype structure has not been investigated at the genetic level. In this study, we have characterized three different Ig heavy chain isotypes of the African lungfish, Protopterus aethiopicus, including an IgM-type heavy chain and short and long forms of non-IgM heavy chains. Northern blot analysis as well as patterns of V(H) utilization suggest that the IgM and non-IgM isotypes are likely encoded in separate loci. The two non-IgM isotypes identified in Protopterus share structural features with the short and long forms of IgX/W/NARC (referred to hereafter as IgW), which were previously considered to be restricted to the cartilaginous fish. It seems that the IgW isotype has a far broader phylogenetic distribution than considered originally and raises questions with regard to the origin and evolutionary divergence of IgM and IgW. Moreover, its absence in other gnathostome lineages implies paradoxically that the IgW-type genes were lost from teleost and tetrapod lineages.
Figures


References
-
- Carroll R L. Vertebrate Paleontology and Evolution. New York: Freeman; 1988.
-
- Helfman G S, Collette B B, Facey D E. The Diversity of Fishes. Oxford: Blackwell; 1997.
-
- Meyer A, Wilson A C. J Mol Evol. 1990;31:359–364. - PubMed
-
- Meyer A, Dolven S I. J Mol Evol. 1992;35:102–113. - PubMed
-
- Marshall C, Schultze H P. J Mol Evol. 1992;35:93–101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
