

Neurovirulence in mice of H5N1 influenza virus genotypes isolated from Hong Kong poultry in 2001
- PMID: 12610156
- PMCID: PMC149508
- DOI: 10.1128/jvi.77.6.3816-3823.2003
Neurovirulence in mice of H5N1 influenza virus genotypes isolated from Hong Kong poultry in 2001
Abstract
We studied the pathogenicity of five different genotypes (A to E) of highly pathogenic avian H5N1 viruses, which contained HA genes similar to those of the H5N1 virus A/goose/Guangdong/1/96 and five different combinations of "internal" genes, in a mouse model. Highly pathogenic, neurotropic variants of genotypes A, C, D, and E were isolated from the brain after a single intranasal passage in mice. Genotype B virus was isolated from lungs only. The mouse brain variants had amino acid changes in all gene products except PB1, NP, and NS1 proteins but no common sets of mutations. We conclude that the original H5N1/01 isolates of genotypes A, C, D, and E were heterogeneous and that highly pathogenic neurotropic variants can be rapidly selected in mice.
Figures
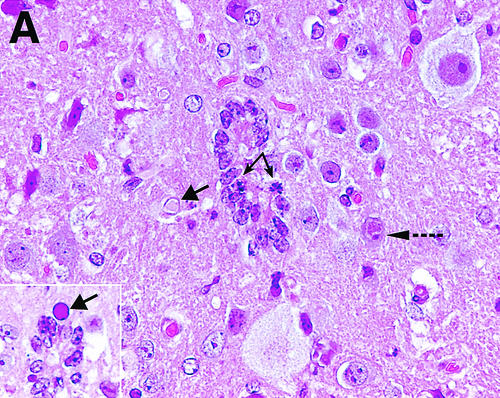
References
-
- Brown, E. G., and J. E. Bailly. 1999. Genetic analysis of mouse-adapted influenza A virus identifies roles for the NA, PB1, and PB2 genes in virulence. Virus Res. 61:63-76. - PubMed
-
- Claas, E. C., A. D. Osterhaus, R. van Beek, J. C. de Jong, G. F. Rimmelzwaan, D. A. Senne, S. Krauss, K. F. Shortridge, and R. G. Webster. 1998. Human influenza A H5N1 virus related to a highly pathogenic avian influenza virus. Lancet 351:472-477. - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
