

GABAA receptor antagonism at the hypoglossal motor nucleus increases genioglossus muscle activity in NREM but not REM sleep
- PMID: 12611924
- PMCID: PMC2342861
- DOI: 10.1113/jphysiol.2002.033696
GABAA receptor antagonism at the hypoglossal motor nucleus increases genioglossus muscle activity in NREM but not REM sleep
Abstract
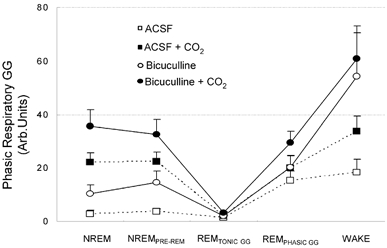
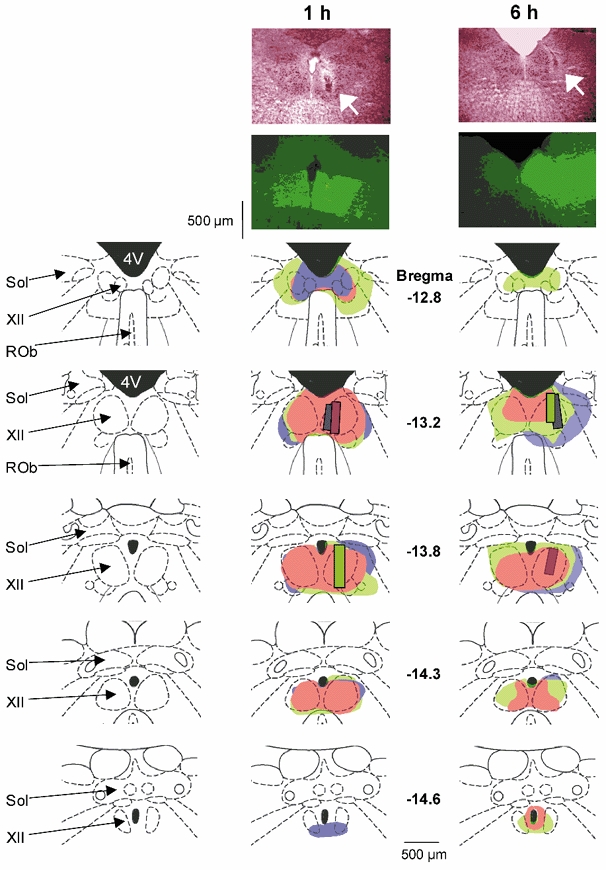
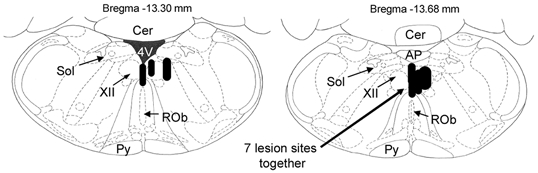
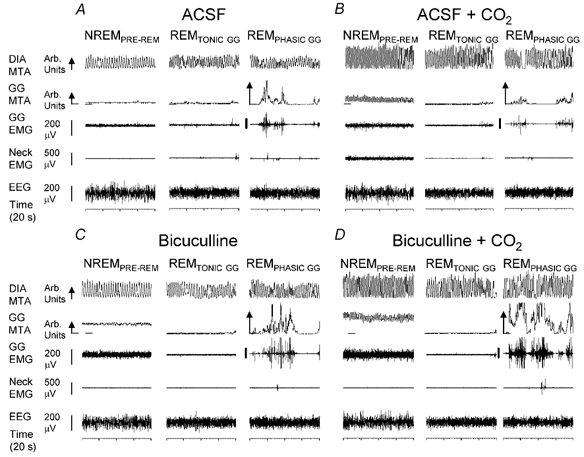
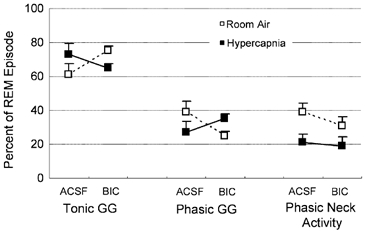
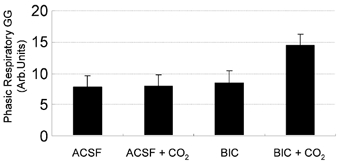
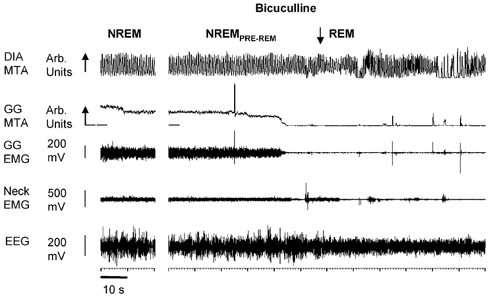
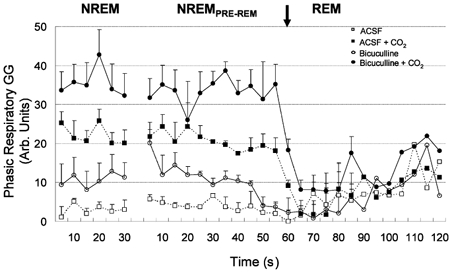
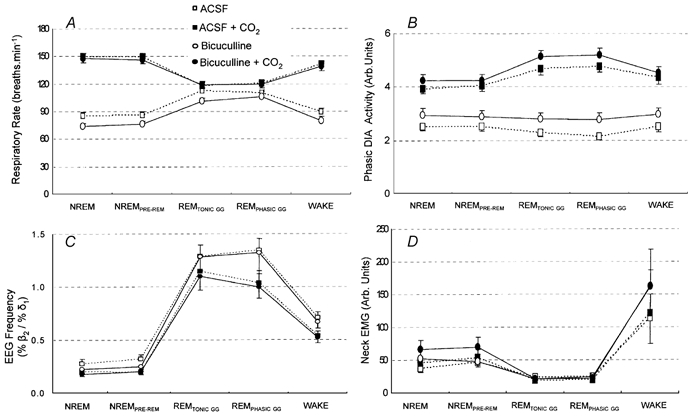
The pharyngeal muscles, such as the genioglossus (GG) muscle of the tongue, are important for effective lung ventilation since they maintain an open airspace. Rapid-eye-movement (REM) sleep, however, recruits powerful neural mechanisms that can abolish GG activity, even during strong reflex respiratory stimulation by elevated CO2. In vitro studies have demonstrated the presence of GABAA receptors on hypoglossal motoneurons, and these and other data have led to the speculation that GABAA mechanisms may contribute to the suppression of hypoglossal motor outflow to the GG muscle in REM sleep. We have developed an animal model that allows us to chronically manipulate neurotransmission at the hypoglossal motor nucleus using microdialysis across natural sleep-wake states in rats. The present study tests the hypothesis that microdialysis perfusion of the GABAA receptor antagonist bicuculline into the hypoglossal motor nucleus will prevent the suppression of GG muscle activity in REM sleep during both room-air and CO2-stimulated breathing. Ten rats were implanted with electroencephalogram and neck muscle electrodes to record sleep-wake states, and GG and diaphragm electrodes for respiratory muscle recording. Microdialysis probes were implanted into the hypoglossal motor nucleus for perfusion of artificial cerebrospinal fluid (ACSF) or 100 microM bicuculline during room-air and CO2-stimulated breathing (7 % inspired CO2). GABAA receptor antagonism at the hypoglossal motor nucleus increased respiratory-related GG activity during both room-air (P = 0.01) and CO2-stimulated breathing (P = 0.007), indicating a background inhibitory GABA tone. However, the effects of bicuculline on GG activity depended on the prevailing sleep-wake state (P < 0.005), with bicuculline increasing GG activity in non-REM (NREM) sleep and wakefulness both in room air and hypercapnia (P < 0.01), but GG activity was effectively abolished in those REM periods without phasic twitches in the GG muscle. This abolition of GG activity in REM sleep occurred regardless of ACSF or bicuculline at the hypoglossal motor nucleus, or room-air or CO2-stimulated breathing (P > 0.63). We conclude that these data in freely behaving rats confirm previous in vitro studies that GABAA receptor mechanisms are present at the hypoglossal motor nucleus and are tonically active, but the data also show that GABAA receptor antagonism at the hypoglossal motor nucleus does not increase GG muscle activity in natural REM sleep.
Figures
Similar articles
-
Role of inhibitory amino acids in control of hypoglossal motor outflow to genioglossus muscle in naturally sleeping rats.J Physiol. 2003 Nov 1;552(Pt 3):975-91. doi: 10.1113/jphysiol.2003.052357. Epub 2003 Aug 22. J Physiol. 2003. PMID: 12937280 Free PMC article.
-
Microdialysis perfusion of 5-HT into hypoglossal motor nucleus differentially modulates genioglossus activity across natural sleep-wake states in rats.J Physiol. 2001 Apr 15;532(Pt 2):467-81. doi: 10.1111/j.1469-7793.2001.0467f.x. J Physiol. 2001. PMID: 11306665 Free PMC article.
-
Suppression of genioglossus muscle tone and activity during reflex hypercapnic stimulation by GABA(A) mechanisms at the hypoglossal motor nucleus in vivo.Neuroscience. 2003;116(1):249-59. doi: 10.1016/s0306-4522(02)00564-x. Neuroscience. 2003. PMID: 12535957
-
K+ channel modulation causes genioglossus inhibition in REM sleep and is a strategy for reactivation.Respir Physiol Neurobiol. 2013 Sep 15;188(3):277-88. doi: 10.1016/j.resp.2013.07.011. Epub 2013 Jul 18. Respir Physiol Neurobiol. 2013. PMID: 23872455 Review.
-
[Selective stimulations and lesions of the rat brain nuclei as the models for research of the human sleep pathology mechanisms].Glas Srp Akad Nauka Med. 2011;(51):85-97. Glas Srp Akad Nauka Med. 2011. PMID: 22165729 Review. Serbian.
Cited by
-
Control of sleep and wakefulness.Physiol Rev. 2012 Jul;92(3):1087-187. doi: 10.1152/physrev.00032.2011. Physiol Rev. 2012. PMID: 22811426 Free PMC article. Review.
-
Neural Control of the Upper Airway: Respiratory and State-Dependent Mechanisms.Compr Physiol. 2016 Sep 15;6(4):1801-1850. doi: 10.1002/cphy.c160002. Compr Physiol. 2016. PMID: 27783860 Free PMC article. Review.
-
The effect of temazepam on assessment of severity of obstructive sleep apnea by polysomnography.Sleep Breath. 2019 Mar;23(1):49-56. doi: 10.1007/s11325-018-1653-3. Epub 2018 Mar 19. Sleep Breath. 2019. PMID: 29556920
-
The temporal structure of REM sleep shows minute-scale fluctuations across brain and body in mice and humans.Proc Natl Acad Sci U S A. 2023 May 2;120(18):e2213438120. doi: 10.1073/pnas.2213438120. Epub 2023 Apr 24. Proc Natl Acad Sci U S A. 2023. PMID: 37094161 Free PMC article.
-
Altered 5-HT2A/C receptor binding in the medulla oblongata in the sudden infant death syndrome (SIDS): Part II. Age-associated alterations in serotonin receptor binding profiles within medullary nuclei supporting cardiorespiratory homeostasis.J Neuropathol Exp Neurol. 2024 Feb 21;83(3):144-160. doi: 10.1093/jnen/nlae004. J Neuropathol Exp Neurol. 2024. PMID: 38323418 Free PMC article.
References
-
- Berger AJ. Determinants of respiratory motoneuron output. Respir Physiol. 2000;122:259–269. - PubMed
-
- Chase MH, Morales FR. Subthreshold excitatory activity and motoneuron discharge during REM periods of active sleep. Science. 1983;221:1195–1198. - PubMed
-
- Chase MH, Morales FR. The control of motoneurons during sleep. In: Kryger MH, Roth T, Dement WC, editors. Principles and Practice of Sleep Medicine. Philadelphia: Blackwell Science Inc; 1994. pp. 163–175.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources