

Roles for SR proteins and hnRNP A1 in the regulation of c-src exon N1
- PMID: 12612063
- PMCID: PMC149473
- DOI: 10.1128/MCB.23.6.1874-1884.2003
Roles for SR proteins and hnRNP A1 in the regulation of c-src exon N1
Abstract
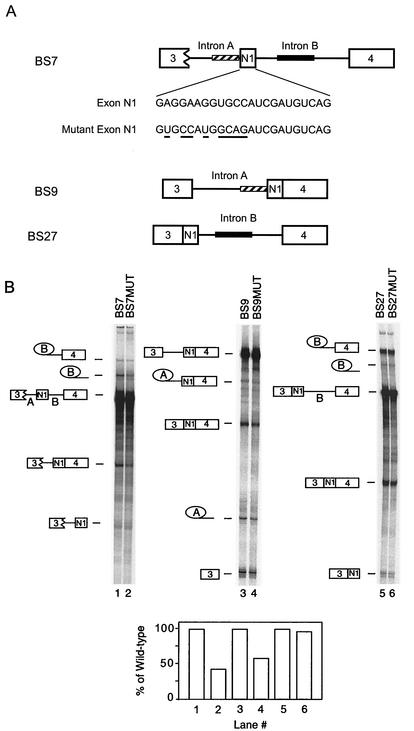
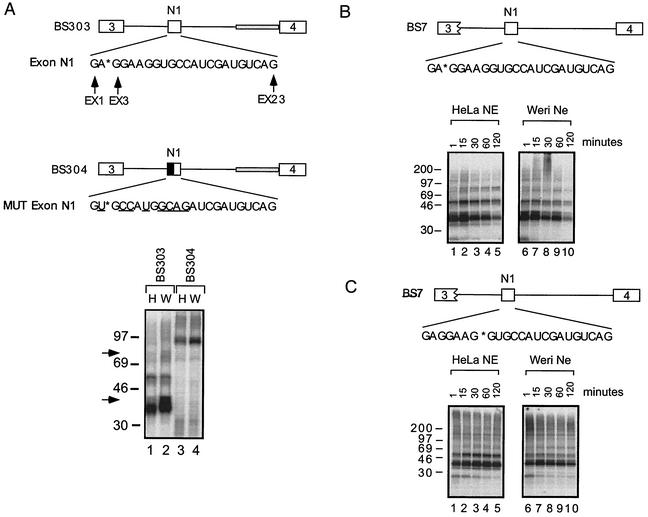
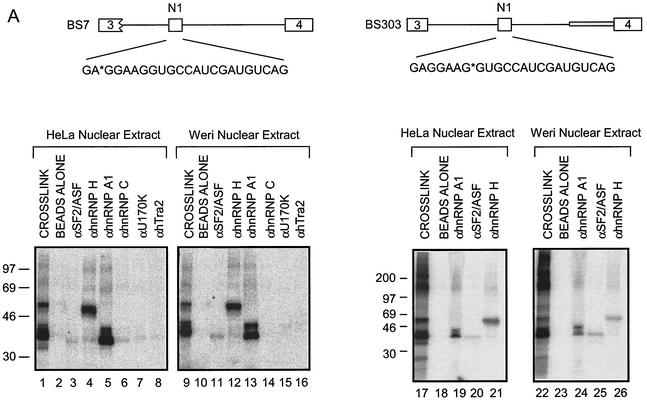
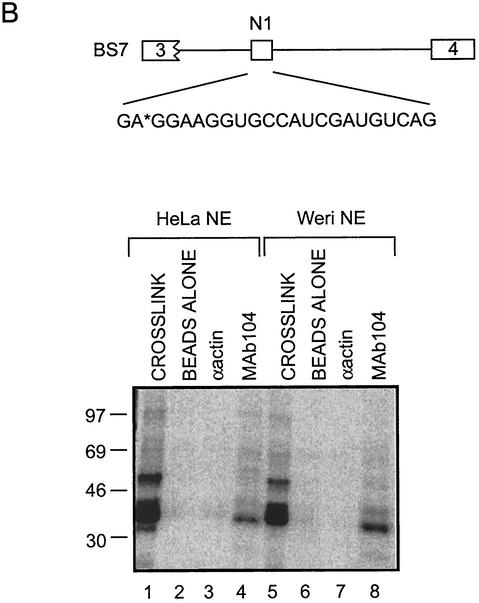
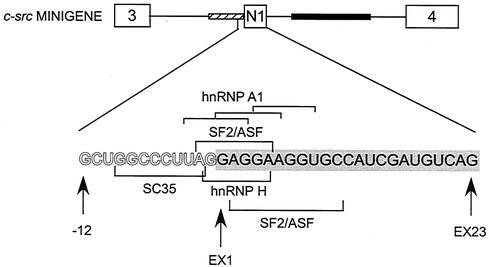
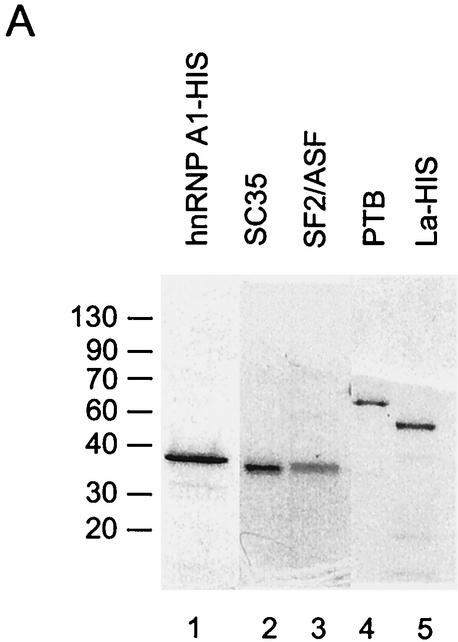
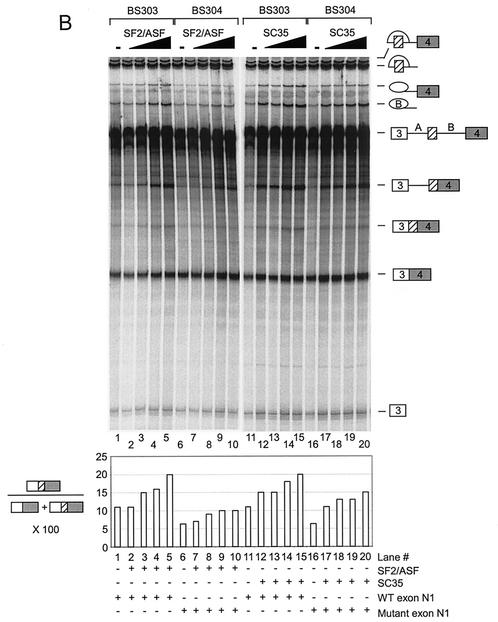
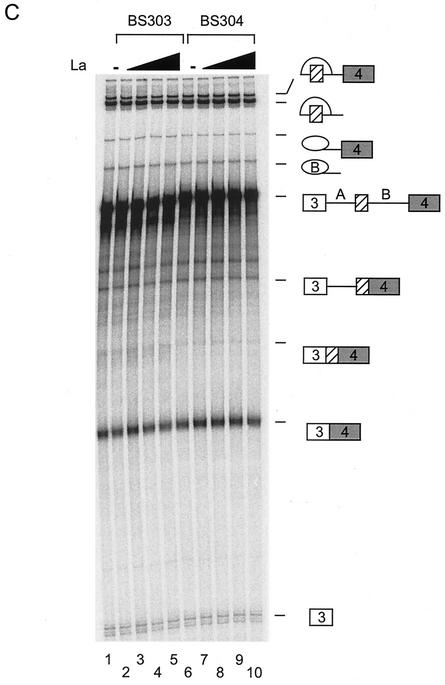
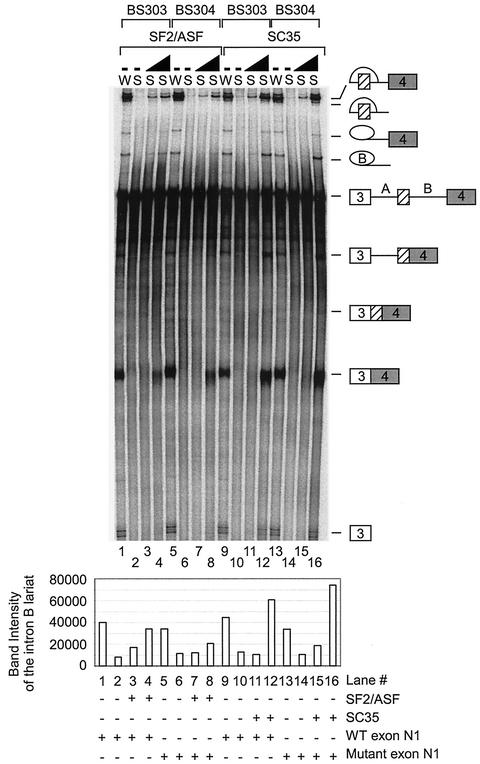
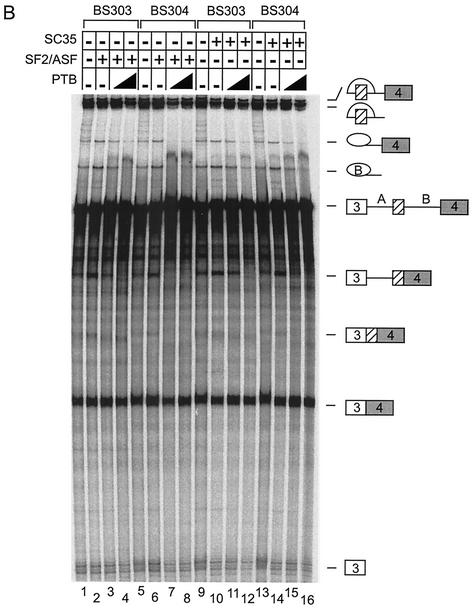
The splicing of the c-src exon N1 is controlled by an intricate combination of positive and negative RNA elements. Most previous work on these sequences focused on intronic elements found upstream and downstream of exon N1. However, it was demonstrated that the 5' half of the N1 exon itself acts as a splicing enhancer in vivo. Here we examine the function of this regulatory element in vitro. We show that a mutation in this sequence decreases splicing of the N1 exon in vitro. Proteins binding to this element were identified as hnRNP A1, hnRNP H, hnRNP F, and SF2/ASF by site-specific cross-linking and immunoprecipitation. The binding of these proteins to the RNA was eliminated by a mutation in the exonic element. The activities of hnRNP A1 and SF2/ASF on N1 splicing were examined by adding purified protein to in vitro splicing reactions. SF2/ASF and another SR protein, SC35, are both able to stimulate splicing of c-src pre-mRNA. However, splicing activation by SF2/ASF is dependent on the N1 exon enhancer element whereas activation by SC35 is not. In contrast to SF2/ASF and in agreement with other systems, hnRNP A1 repressed c-src splicing in vitro. The negative activity of hnRNP A1 on splicing was compared with that of PTB, a protein previously demonstrated to repress splicing in this system. Both proteins repress exon N1 splicing, and both counteract the enhancing activity of the SR proteins. Removal of the PTB binding sites upstream of N1 prevents PTB-mediated repression but does not affect A1-mediated repression. Thus, hnRNP A1 and PTB use different mechanisms to repress c-src splicing. Our results link the activity of these well-known exonic splicing regulators, SF2/ASF and hnRNP A1, to the splicing of an exon primarily controlled by intronic factors.
Figures



Similar articles
-
hnRNP A1 and the SR proteins ASF/SF2 and SC35 have antagonistic functions in splicing of beta-tropomyosin exon 6B.J Biol Chem. 2004 Sep 10;279(37):38249-59. doi: 10.1074/jbc.M405377200. Epub 2004 Jun 18. J Biol Chem. 2004. PMID: 15208309
-
Exon identity established through differential antagonism between exonic splicing silencer-bound hnRNP A1 and enhancer-bound SR proteins.Mol Cell. 2001 Dec;8(6):1351-61. doi: 10.1016/s1097-2765(01)00409-9. Mol Cell. 2001. PMID: 11779509
-
Modulation of exon skipping and inclusion by heterogeneous nuclear ribonucleoprotein A1 and pre-mRNA splicing factor SF2/ASF.Mol Cell Biol. 1993 May;13(5):2993-3001. doi: 10.1128/mcb.13.5.2993-3001.1993. Mol Cell Biol. 1993. PMID: 8474457 Free PMC article.
-
Functional interactions between polypyrimidine tract binding protein and PRI peptide ligand containing proteins.Biochem Soc Trans. 2016 Aug 15;44(4):1058-65. doi: 10.1042/BST20160080. Biochem Soc Trans. 2016. PMID: 27528752 Review.
-
Evolution of SR protein and hnRNP splicing regulatory factors.Wiley Interdiscip Rev RNA. 2012 Jan-Feb;3(1):1-12. doi: 10.1002/wrna.100. Epub 2011 Sep 2. Wiley Interdiscip Rev RNA. 2012. PMID: 21898828 Free PMC article. Review.
Cited by
-
Exon inclusion is dependent on predictable exonic splicing enhancers.Mol Cell Biol. 2005 Aug;25(16):7323-32. doi: 10.1128/MCB.25.16.7323-7332.2005. Mol Cell Biol. 2005. PMID: 16055740 Free PMC article.
-
hnRNP A1 Regulates Alternative Splicing of Tau Exon 10 by Targeting 3' Splice Sites.Cells. 2020 Apr 10;9(4):936. doi: 10.3390/cells9040936. Cells. 2020. PMID: 32290247 Free PMC article.
-
Elevated copper remodels hepatic RNA processing machinery in the mouse model of Wilson's disease.J Mol Biol. 2011 Feb 11;406(1):44-58. doi: 10.1016/j.jmb.2010.12.001. Epub 2010 Dec 10. J Mol Biol. 2011. PMID: 21146535 Free PMC article.
-
RNA-Binding Proteins: Splicing Factors and Disease.Biomolecules. 2015 May 13;5(2):893-909. doi: 10.3390/biom5020893. Biomolecules. 2015. PMID: 25985083 Free PMC article. Review.
-
Diverse regulation of 3' splice site usage.Cell Mol Life Sci. 2015 Dec;72(24):4771-93. doi: 10.1007/s00018-015-2037-5. Epub 2015 Sep 14. Cell Mol Life Sci. 2015. PMID: 26370726 Free PMC article. Review.
References
-
- Black, D. L. 1992. Activation of c-src neuron-specific splicing by an unusual RNA element in vivo and in vitro. Cell 69:795-807. - PubMed
-
- Black, D. L. 1991. Does steric interference between splice sites block the splicing of a short c-src neuron-specific exon in non-neuronal cells? Genes Dev. 5:389-402. - PubMed
-
- Blencowe, B. J. 2000. Exonic splicing enhancers: mechanism of action, diversity and role in human genetic diseases. Trends Biochem. Sci. 25:106-110. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous