

Regulation of cytochrome c oxidase activity by c-Src in osteoclasts
- PMID: 12615910
- PMCID: PMC2173369
- DOI: 10.1083/jcb.200209098
Regulation of cytochrome c oxidase activity by c-Src in osteoclasts
Abstract
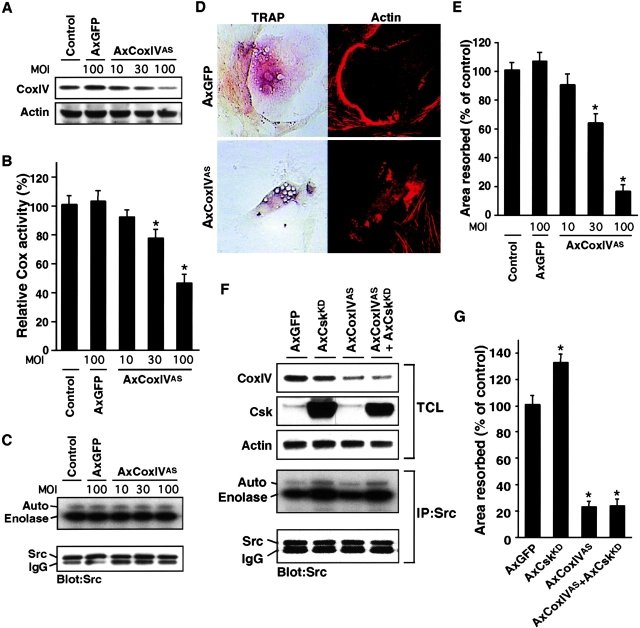
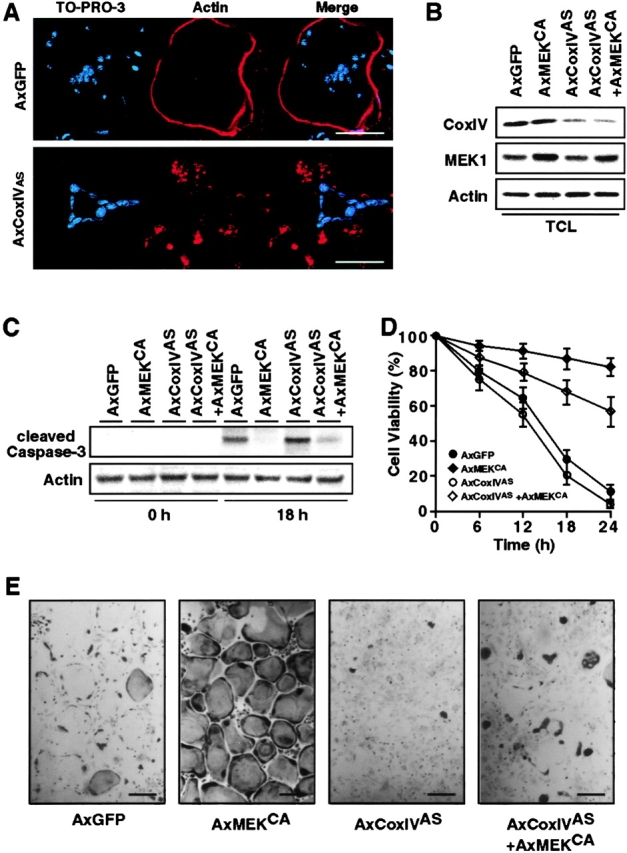
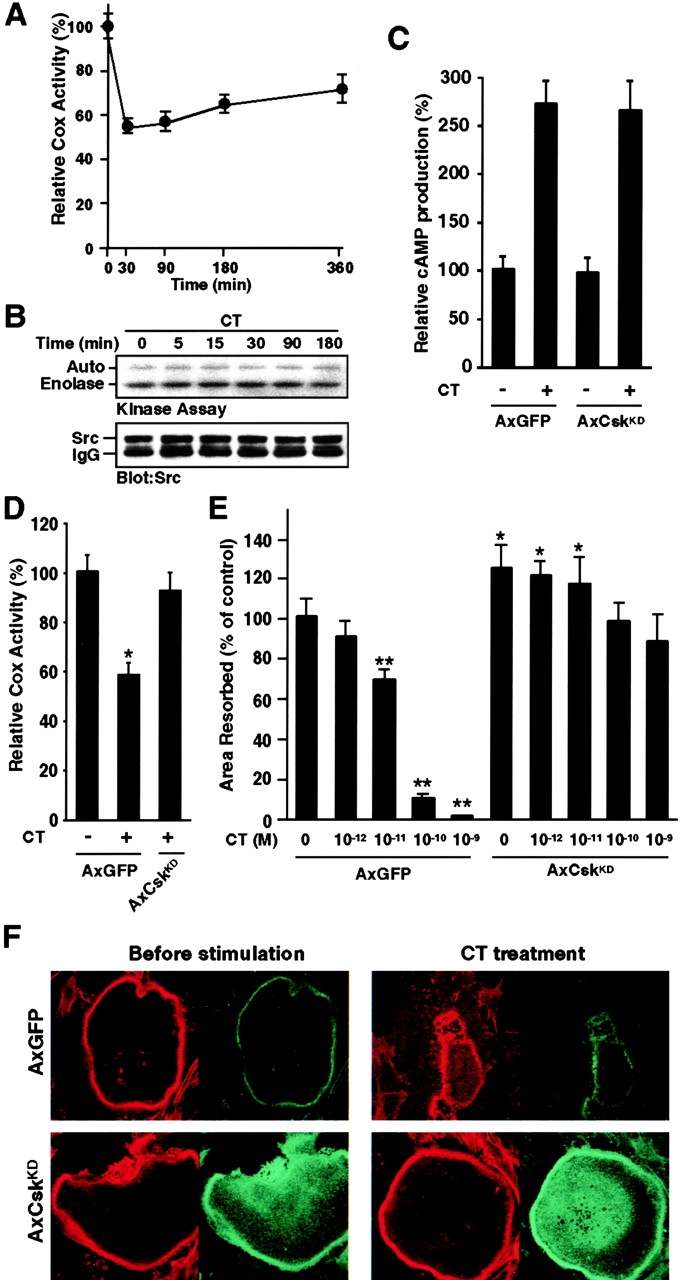
The function of the nonreceptor tyrosine kinase c-Src as a plasma membrane-associated molecular effector of a variety of extracellular stimuli is well known. Here, we show that c-Src is also present within mitochondria, where it phosphorylates cytochrome c oxidase (Cox). Deleting the c-src gene reduces Cox activity, and this inhibitory effect is restored by expressing exogenous c-Src. Furthermore, reducing endogenous Src kinase activity down-regulates Cox activity, whereas activating Src has the opposite effect. Src-induced Cox activity is required for normal function of cells that require high levels of ATP, such as mitochondria-rich osteoclasts. The peptide hormone calcitonin, which inhibits osteoclast function, also down-regulates Cox activity. Increasing Src kinase activity prevented the inhibitory effect of calcitonin on Cox activity and osteoclast function. These results suggest that c-Src plays a previously unrecognized role in maintaining cellular energy stores by activating Cox in mitochondria.
Figures
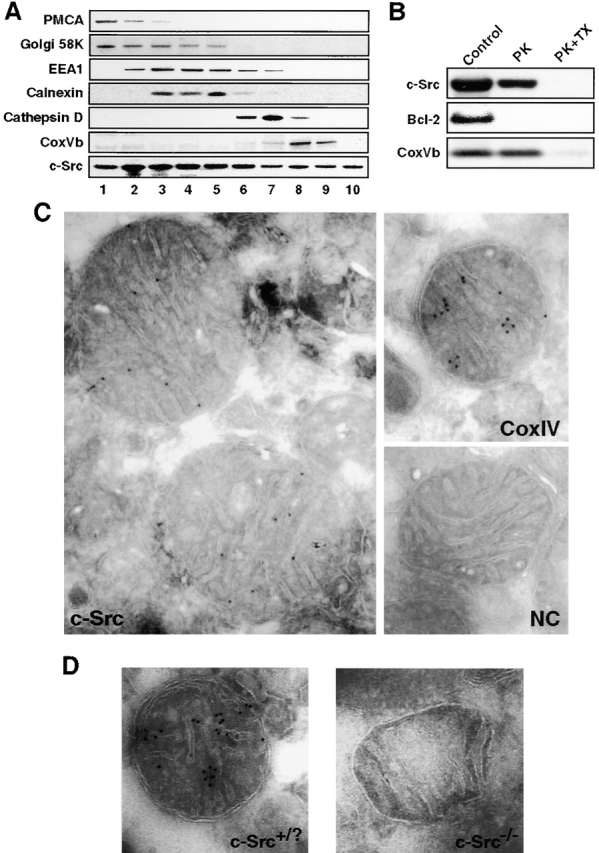
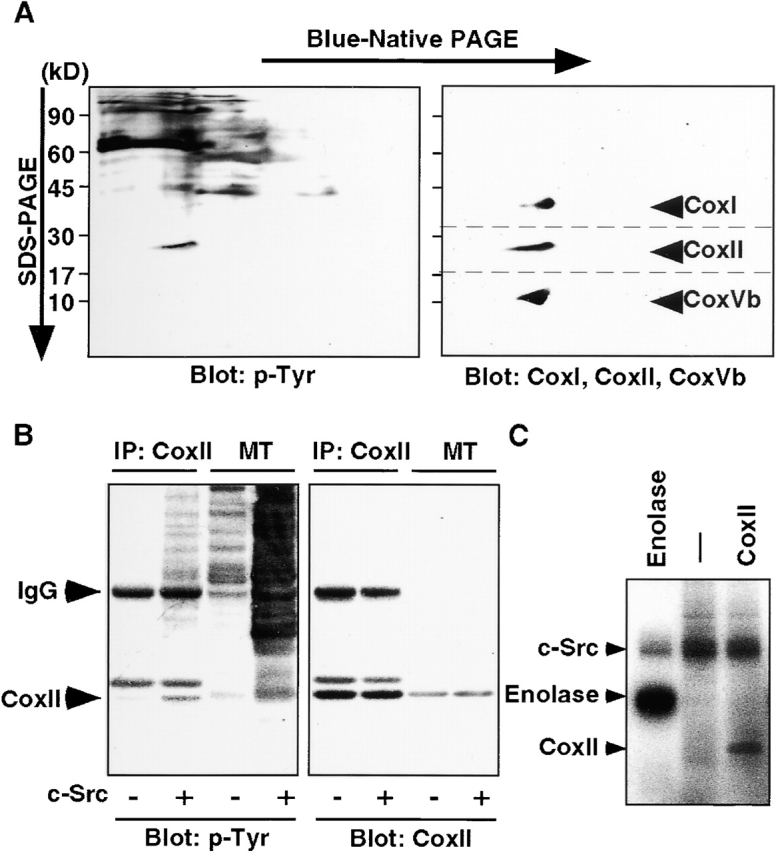
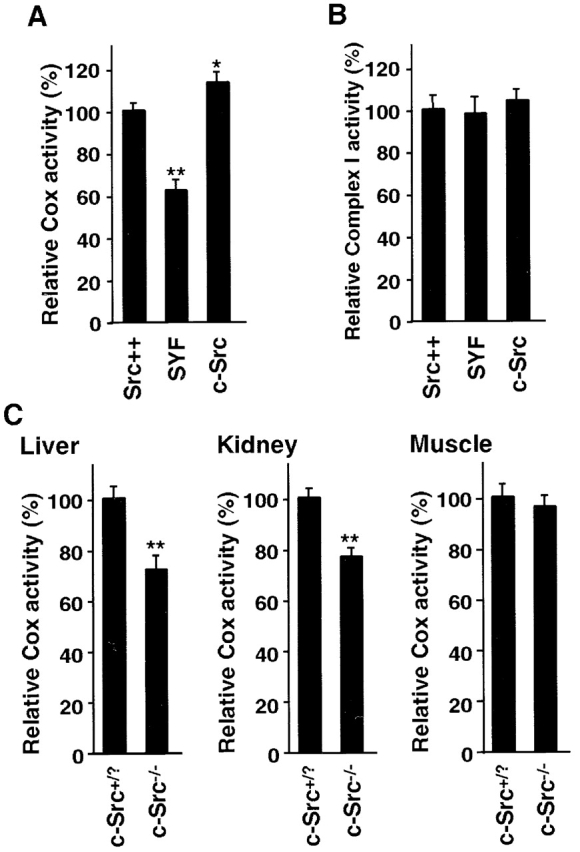
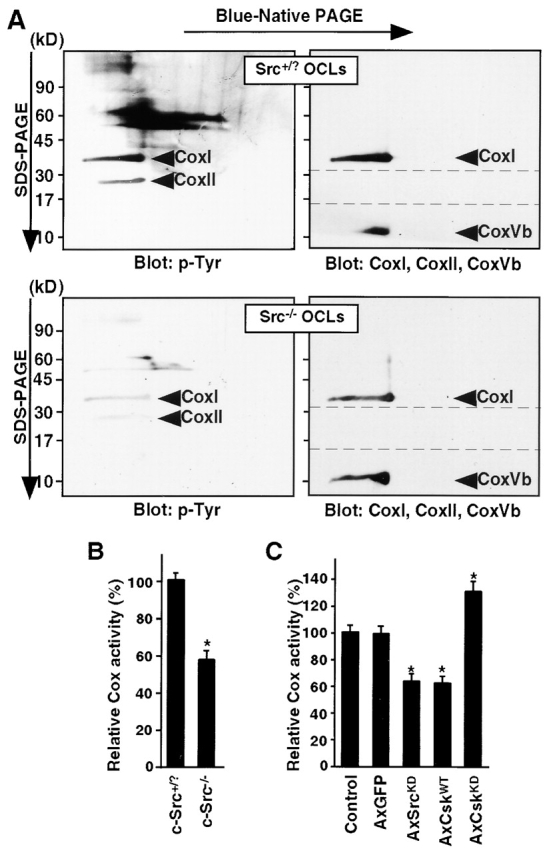
References
-
- Bard, F., U. Patel, J.B. Levy, W.C. Horne, and R. Baron. 2002. Molecular complexes that contain both c-Cbl and c-Src associate with Golgi membranes. Eur. J. Cell Biol. 81:26–35. - PubMed
-
- Baron, R., J.-H. Ravesloot, L. Neff, M. Chakraborty, D. Chatterjee, A. Lomri, and W. Horne. 1993. Cellular and molecular biology of the osteoclast. Cellular and Molecular Biology of Bone. M. Noda, editor. Academic Press, San Diego. 445–495.
-
- Brown, M.T., and J.A. Cooper. 1996. Regulation, substrates and functions of src. Biochim. Biophys. Acta. 1287:121–149. - PubMed
-
- Burke, P.A., and R.O. Poyton. 1998. Structure/function of oxygen-regulated isoforms in cytochrome c oxidase. J. Exp. Biol. 201:1163–1175. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
