

Cytokinesis in bacteria
- PMID: 12626683
- PMCID: PMC150516
- DOI: 10.1128/MMBR.67.1.52-65.2003
Cytokinesis in bacteria
Abstract
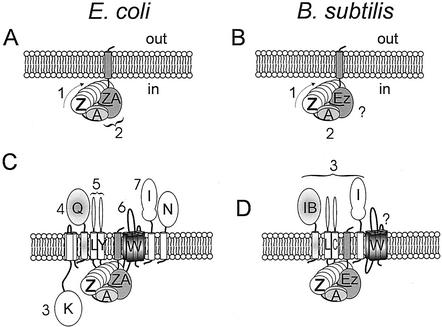
Work on two diverse rod-shaped bacteria, Escherichia coli and Bacillus subtilis, has defined a set of about 10 conserved proteins that are important for cell division in a wide range of eubacteria. These proteins are directed to the division site by the combination of two negative regulatory systems. Nucleoid occlusion is a poorly understood mechanism whereby the nucleoid prevents division in the cylindrical part of the cell, until chromosome segregation has occurred near midcell. The Min proteins prevent division in the nucleoid-free spaces near the cell poles in a manner that is beginning to be understood in cytological and biochemical terms. The hierarchy whereby the essential division proteins assemble at the midcell division site has been worked out for both E. coli and B. subtilis. They can be divided into essentially three classes depending on their position in the hierarchy and, to a certain extent, their subcellular localization. FtsZ is a cytosolic tubulin-like protein that polymerizes into an oligomeric structure that forms the initial ring at midcell. FtsA is another cytosolic protein that is related to actin, but its precise function is unclear. The cytoplasmic proteins are linked to the membrane by putative membrane anchor proteins, such as ZipA of E. coli and possibly EzrA of B. subtilis, which have a single membrane span but a cytoplasmic C-terminal domain. The remaining proteins are either integral membrane proteins or transmembrane proteins with their major domains outside the cell. The functions of most of these proteins are unclear with the exception of at least one penicillin-binding protein, which catalyzes a key step in cell wall synthesis in the division septum.
Figures

References
-
- Addinall, S. G., C. Cao, and J. Lutkenhaus. 1997. FtsN, a late recruit to the septum in Escherichia coli. Mol. Microbiol. 25:303-309. - PubMed
-
- Addinall, S. G., C. Cao, and J. Lutkenhaus. 1997. Temperature shift experiments with an ftsZ84(Ts) strain reveal rapid dynamics of FtsZ localization and indicate that the Z ring is required throughout septation and cannot reoccupy division sites once constriction has initiated. J. Bacteriol. 179:4277-4284. - PMC - PubMed
-
- Addinall, S. G., and J. Lutkenhaus. 1996. FtsZ-spirals and -arcs determine the shape of the invaginating septa in some mutants of Escherichia coli. Mol. Microbiol. 22:231-237. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
