

Bacteriophage T4 genome
- PMID: 12626685
- PMCID: PMC150520
- DOI: 10.1128/MMBR.67.1.86-156.2003
Bacteriophage T4 genome
Abstract
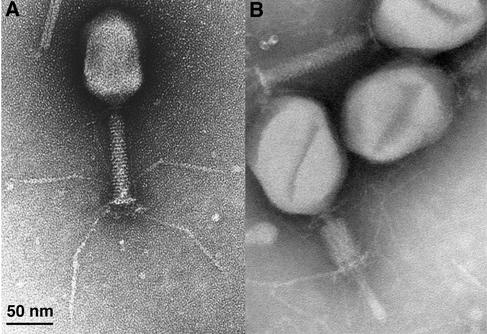
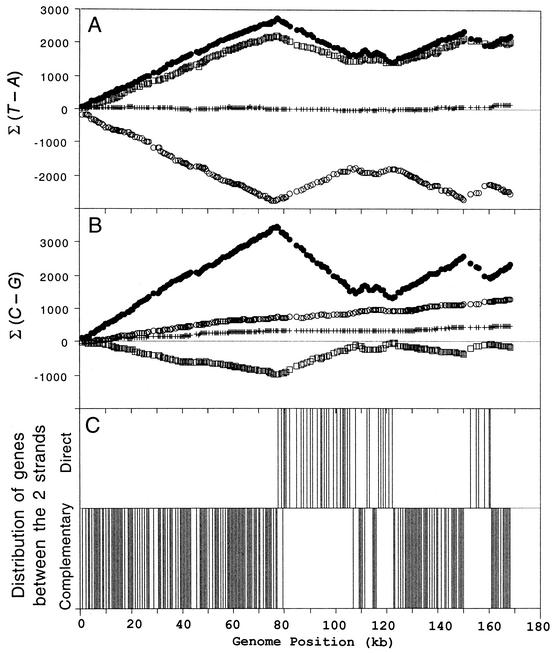
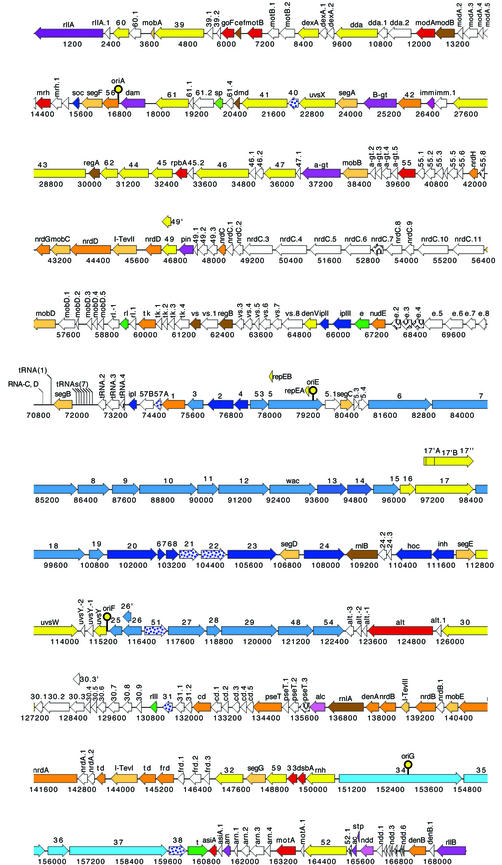
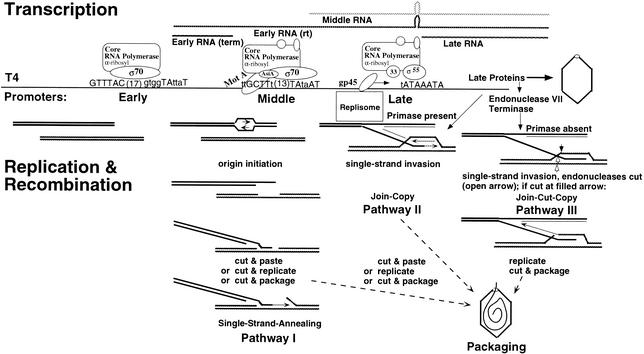
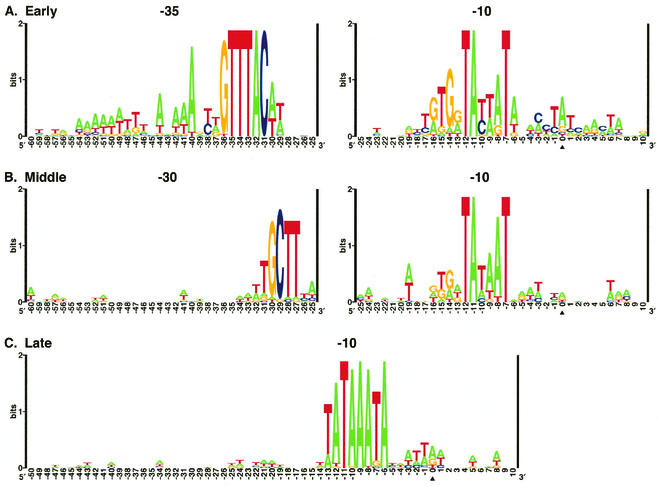
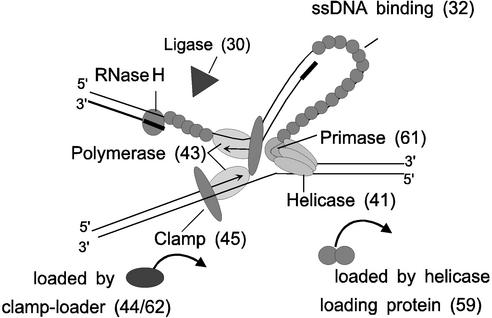
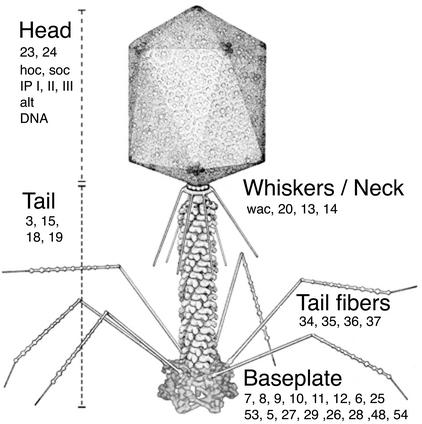
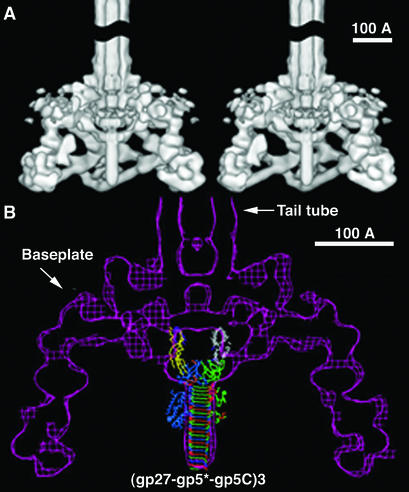
Phage T4 has provided countless contributions to the paradigms of genetics and biochemistry. Its complete genome sequence of 168,903 bp encodes about 300 gene products. T4 biology and its genomic sequence provide the best-understood model for modern functional genomics and proteomics. Variations on gene expression, including overlapping genes, internal translation initiation, spliced genes, translational bypassing, and RNA processing, alert us to the caveats of purely computational methods. The T4 transcriptional pattern reflects its dependence on the host RNA polymerase and the use of phage-encoded proteins that sequentially modify RNA polymerase; transcriptional activator proteins, a phage sigma factor, anti-sigma, and sigma decoy proteins also act to specify early, middle, and late promoter recognition. Posttranscriptional controls by T4 provide excellent systems for the study of RNA-dependent processes, particularly at the structural level. The redundancy of DNA replication and recombination systems of T4 reveals how phage and other genomes are stably replicated and repaired in different environments, providing insight into genome evolution and adaptations to new hosts and growth environments. Moreover, genomic sequence analysis has provided new insights into tail fiber variation, lysis, gene duplications, and membrane localization of proteins, while high-resolution structural determination of the "cell-puncturing device," combined with the three-dimensional image reconstruction of the baseplate, has revealed the mechanism of penetration during infection. Despite these advances, nearly 130 potential T4 genes remain uncharacterized. Current phage-sequencing initiatives are now revealing the similarities and differences among members of the T4 family, including those that infect bacteria other than Escherichia coli. T4 functional genomics will aid in the interpretation of these newly sequenced T4-related genomes and in broadening our understanding of the complex evolution and ecology of phages-the most abundant and among the most ancient biological entities on Earth.
Figures

References
-
- Abdus Sattar, A. K., T. C. Lin, C. Jones, and W. H. Konigsberg. 1996. Functional consequences and exonuclease kinetic parameters of point mutations in bacteriophage T4 DNA polymerase. Biochemistry 35:16621-16629. - PubMed
-
- Abedon, S. T. 1999. Bacteriophage T4 resistance to lysis-inhibition collapse. Genet. Res. 74:1-11. - PubMed
-
- Abedon, S. T. 1994. Lysis and the interaction between free phages and infected cells., p. 397-405. In J. Karam, J. W. Drake, K. N. Kreuzer, G. Mosig, D. H. Hall, F. A. Eiserling, L. W. Black, E. K. Spicer, E. Kutter, K. Carlson, and E. S. Miller (ed.), Molecular biology of bacteriophage T4. American Society for Microbiology, Washington, D.C.
-
- Abedon, S. T. 1990. Selection for lysis inhibition in bacteriophage. J. Theor. Biol. 146:501-511. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
