

The complex between a four-way DNA junction and T7 endonuclease I
- PMID: 12628932
- PMCID: PMC151070
- DOI: 10.1093/emboj/cdg132
The complex between a four-way DNA junction and T7 endonuclease I
Abstract
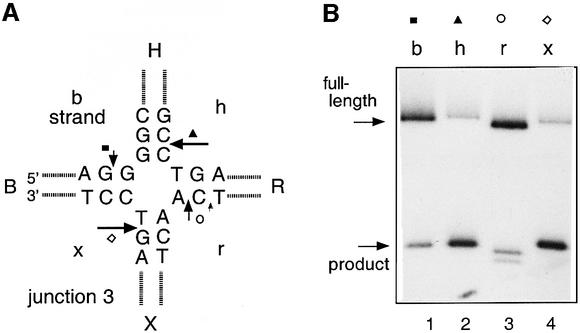
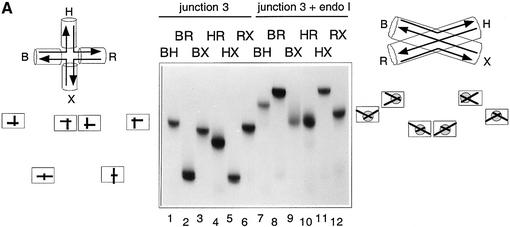
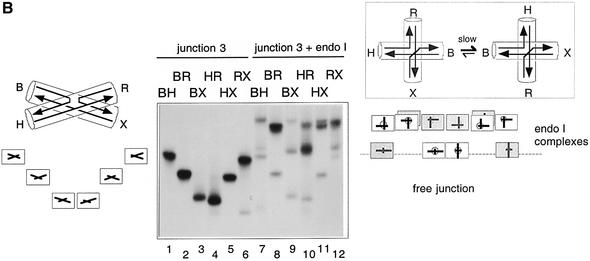
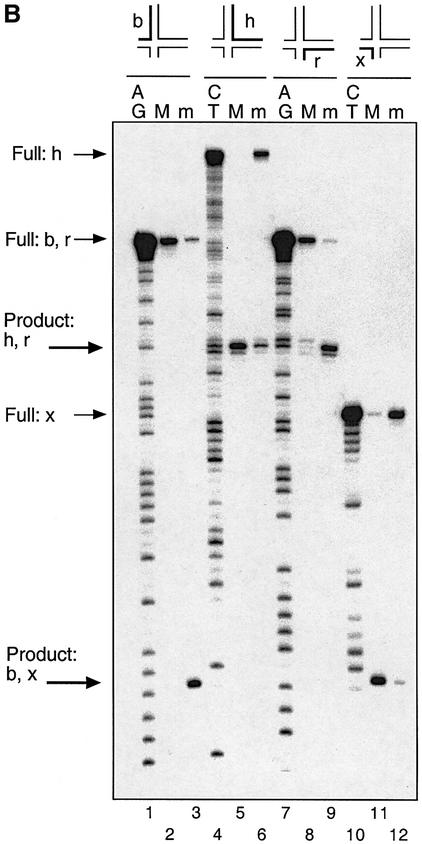
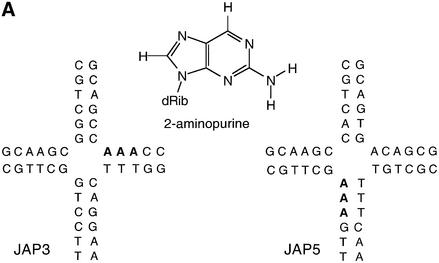
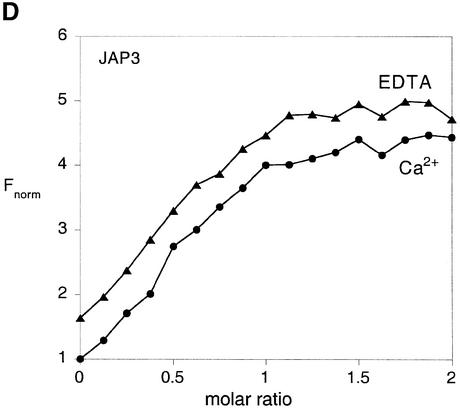
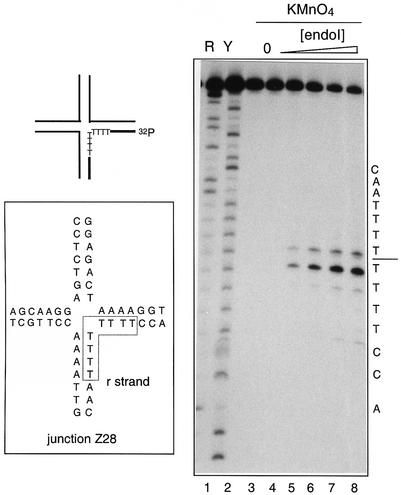
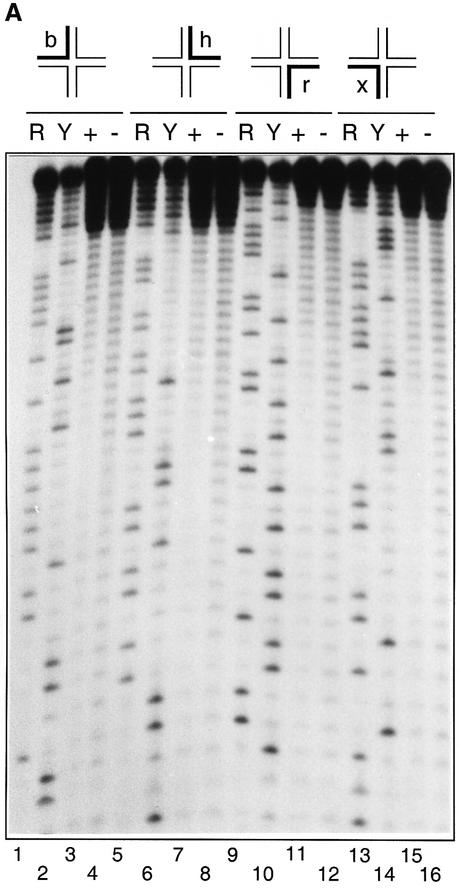
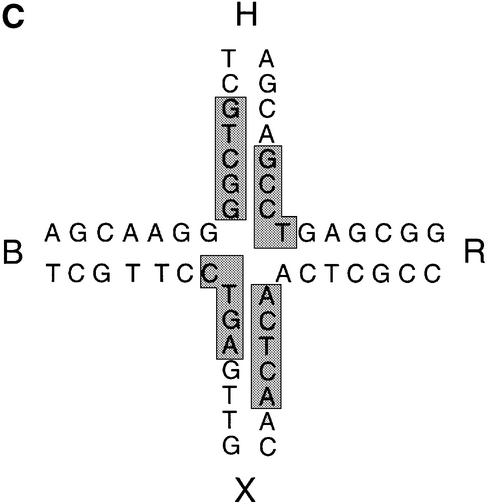
The junction-resolving enzyme endonuclease I is selective for the structure of the DNA four-way (Holliday) junction. The enzyme binds to a four-way junction in two possible orientations, with a 4:1 ratio, opening the DNA structure at the centre and changing the global structure into a 90 degrees cross of approximately coaxial helices. The nuclease cleaves the continuous strands of the junction in each orientation. Binding leads to pronounced regions of protection of the DNA against hydroxyl radical attack. Using all this information together with the known structure of the enzyme and the structure of the BglI-DNA complex, we have constructed a model of the complex of endonuclease I and a DNA junction. This shows how the enzyme is selective for the structure of a four-way junction, such that both continuous strands can be accommodated into the two active sites so that a productive resolution event is possible.
Figures









References
-
- Ariyoshi M., Vassylyev,D.G., Iwasaki,H., Nakamura,H., Shinagawa,H. and Morikawa,K. (1994) Atomic structure of the RuvC resolvase: a Holliday junction-specific endonuclease from E.coli. Cell, 78, 1063–1072. - PubMed
-
- Beaucage S.L. and Caruthers,M.H. (1981) Deoxynucleoside phosphor amidites—a new class of key intermediates for deoxypolynucleotide synthesis. Tetrahedron Lett., 22, 1859–1862.
-
- Bennett R.J. and West,S.C. (1995) Structural analysis of the RuvC–Holliday junction complex reveals an unfolded junction. J. Mol. Biol., 252, 213–226. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
