

Suppression of Ras-stimulated transformation by the JNK signal transduction pathway
- PMID: 12629045
- PMCID: PMC196007
- DOI: 10.1101/gad.1062903
Suppression of Ras-stimulated transformation by the JNK signal transduction pathway
Abstract
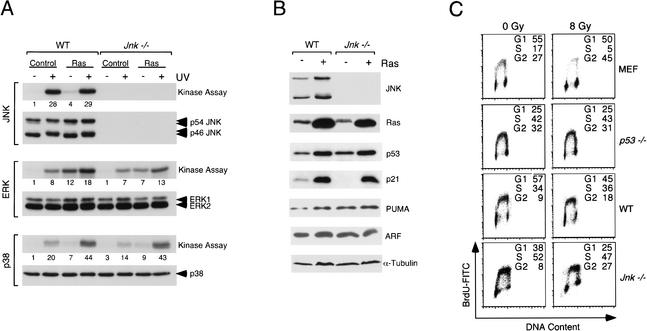
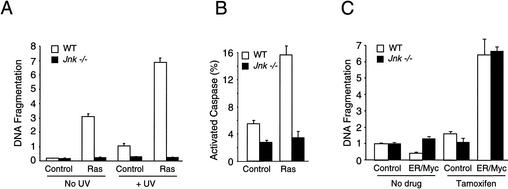
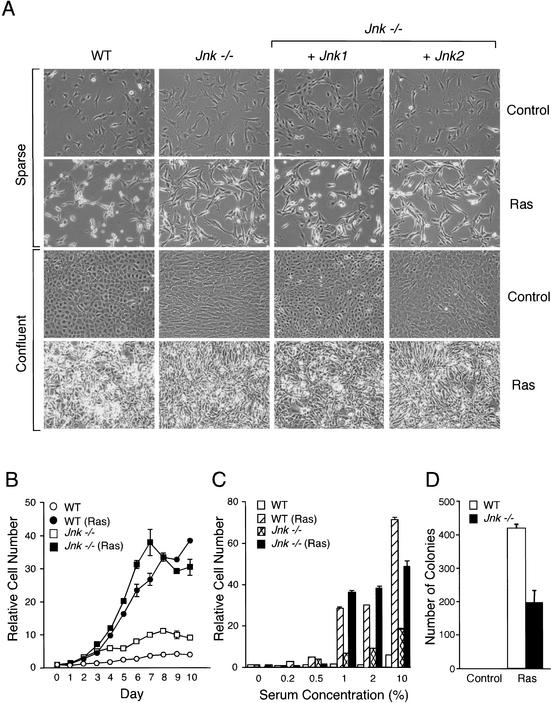
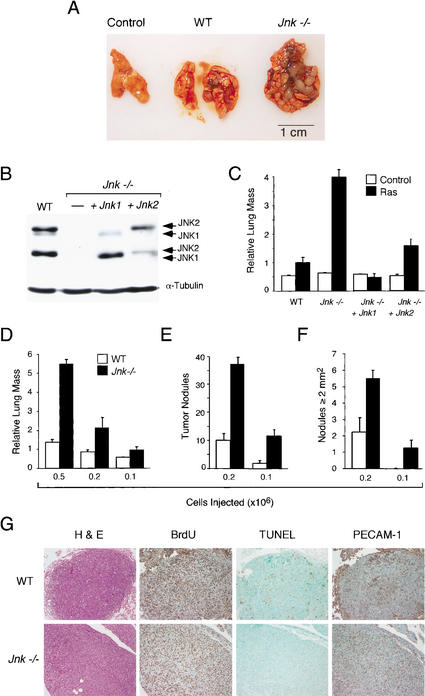
The c-Jun NH(2)-terminal kinase (JNK) phosphorylates and activates members of the activator protein-1 (AP-1) group of transcription factors and is implicated in oncogenic transformation. To examine the role of JNK, we investigated the effect of JNK deficiency on Ras-stimulated transformation. We demonstrate that although JNK does play a role in transformation in vitro, JNK is not required for tumor development in vivo. Importantly, the loss of JNK expression resulted in substantial increases in the number and growth of tumor nodules in vivo. Complementation assays demonstrated that this phenotype was caused by JNK deficiency. These data demonstrate that, in contrast to expectations, the normal function of JNK may be to suppress tumor development in vivo. This conclusion is consistent with the presence in human tumors of loss-of-function mutations in the JNK pathway.
Figures
References
-
- Behrens A, Jochum W, Sibilia M, Wagner EF. Oncogenic transformation by ras and fos is mediated by c-Jun N-terminal phosphorylation. Oncogene. 2000;19:2657–2663. - PubMed
-
- Bos JL. Ras oncogenes in human cancer: A review. Cancer Res. 1989;49:4682–4689. - PubMed
-
- Clark GJ, Cox AD, Graham SM, Der CJ. Biological assays for Ras transformation. Methods Enzymol. 1995;255:395–412. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous