

Neuronal basis of crossed actions from the reticular formation on feline hindlimb motoneurons
- PMID: 12629191
- PMCID: PMC1890022
- DOI: 10.1523/JNEUROSCI.23-05-01867.2003
Neuronal basis of crossed actions from the reticular formation on feline hindlimb motoneurons
Abstract
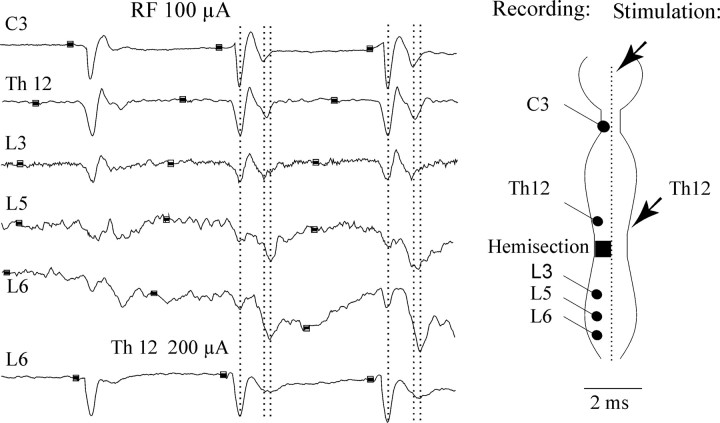
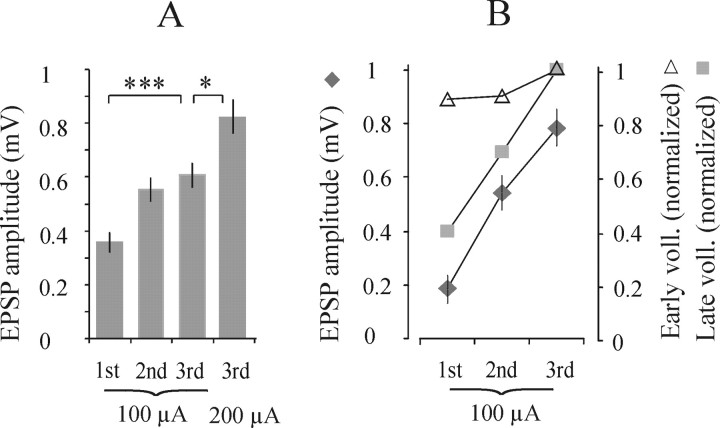
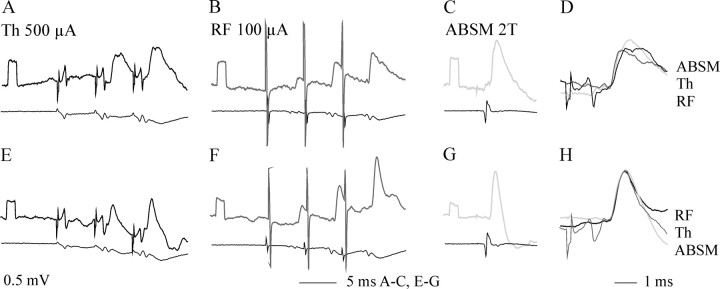
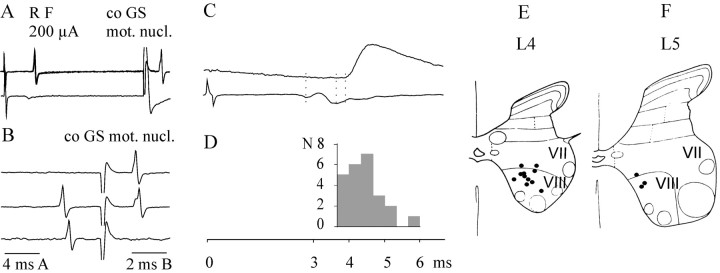
Pathways through which reticulospinal neurons can influence contralateral limb movements were investigated by recording from motoneurons innervating hindlimb muscles. Reticulospinal tract fibers were stimulated within the brainstem or in the lateral funiculus of the thoracic spinal cord contralateral to the motoneurons. Effects evoked by ipsilaterally descending reticulospinal tract fibers were eliminated by a spinal hemisection at an upper lumbar level. Stimuli applied in the brainstem evoked EPSPs, IPSPs, or both at latencies of 1.42 +/- 0.03 and 1.53 +/- 0.04 msec, respectively, from the first components of the descending volleys and with properties indicating a disynaptic linkage, in most contralateral motoneurons: EPSPs in 76% and IPSPs in 26%. EPSPs with characteristics of monosynaptically evoked responses, attributable to direct actions of crossed axon collaterals of reticulospinal fibers, were found in a small proportion of the motoneurons, whether evoked from the brainstem (9%) or from the thoracic cord (12.5%). Commissural neurons, which might mediate the crossed disynaptic actions (i.e., were antidromically activated from contralateral motor nuclei and monosynaptically excited from the ipsilateral reticular formation), were found in Rexed's lamina VIII in the midlumbar segments (L3-L5). The results reveal that although direct actions of reticulospinal fibers are much more potent on ipsilateral motoneurons, interneuronally mediated actions are as potent contralaterally as ipsilaterally, and midlumbar commissural neurons are likely to contribute to them. They indicate a close coupling between the spinal interneuronal systems used by the reticulospinal neurons to coordinate muscle contractions ipsilaterally and contralaterally.
Figures






Similar articles
-
Are crossed actions of reticulospinal and vestibulospinal neurons on feline motoneurons mediated by the same or separate commissural neurons?J Neurosci. 2003 Sep 3;23(22):8041-50. doi: 10.1523/JNEUROSCI.23-22-08041.2003. J Neurosci. 2003. PMID: 12954866 Free PMC article.
-
Uncrossed actions of feline corticospinal tract neurones on hindlimb motoneurones evoked via ipsilaterally descending pathways.J Physiol. 2007 Apr 1;580(Pt 1):119-32. doi: 10.1113/jphysiol.2006.122721. Epub 2007 Jan 25. J Physiol. 2007. PMID: 17255171 Free PMC article.
-
Same spinal interneurons mediate reflex actions of group Ib and group II afferents and crossed reticulospinal actions.J Neurophysiol. 2006 Jun;95(6):3911-22. doi: 10.1152/jn.01262.2005. Epub 2006 Mar 22. J Neurophysiol. 2006. PMID: 16554505 Free PMC article.
-
The spinal motor system in early vertebrates and some of its evolutionary changes.Brain Behav Evol. 1992;40(2-3):82-97. doi: 10.1159/000113905. Brain Behav Evol. 1992. PMID: 1422809 Review.
-
Reticulospinal Systems for Tuning Motor Commands.Front Neural Circuits. 2018 Apr 18;12:30. doi: 10.3389/fncir.2018.00030. eCollection 2018. Front Neural Circuits. 2018. PMID: 29720934 Free PMC article. Review.
Cited by
-
Premotor interneurones contributing to actions of feline pyramidal tract neurones on ipsilateral hindlimb motoneurones.J Physiol. 2008 Jan 15;586(2):557-74. doi: 10.1113/jphysiol.2007.145466. Epub 2007 Nov 15. J Physiol. 2008. PMID: 18006578 Free PMC article.
-
Modulation between bilateral legs and within unilateral muscle synergists of postural muscle activity changes with development and aging.Exp Brain Res. 2014 Jan;232(1):1-11. doi: 10.1007/s00221-013-3702-2. Epub 2013 Nov 10. Exp Brain Res. 2014. PMID: 24240388
-
Propriospinal neurons are sufficient for bulbospinal transmission of the locomotor command signal in the neonatal rat spinal cord.J Physiol. 2008 Mar 15;586(6):1623-35. doi: 10.1113/jphysiol.2007.148361. Epub 2008 Jan 31. J Physiol. 2008. PMID: 18238807 Free PMC article.
-
A trans-spinal loop between neurones in the reticular formation and in the cerebellum.J Physiol. 2011 Feb 1;589(Pt 3):653-65. doi: 10.1113/jphysiol.2010.201178. Epub 2010 Dec 13. J Physiol. 2011. PMID: 21149461 Free PMC article.
-
Ipsilateral actions from the feline red nucleus on hindlimb motoneurones.J Physiol. 2008 Dec 15;586(24):5865-84. doi: 10.1113/jphysiol.2008.163998. Epub 2008 Oct 20. J Physiol. 2008. PMID: 18936076 Free PMC article.
References
-
- Aggelopoulos NC, Edgley SA. Segmental localisation of the relays mediating crossed inhibition of hindlimb motoneurones from group II afferents in the anaesthetized cat spinal cord. Neurosci Lett. 1995;185:60–64. - PubMed
-
- Alstermark B, Kummel H. Transneuronal transport of wheat germ agglutinin conjugated horseradish peroxidase into last order spinal interneurones projecting to acromio- and spinodeltoideus motoneurones in the cat. 1. Location of labelled interneurones and influence of synaptic activity on the transneuronal transport. Exp Brain Res. 1990;80:83–95. - PubMed
-
- Burke RE. Composite nature of the monosynaptic excitatory postsynaptic potential. J Neurophysiol. 1967;30:1114–1137. - PubMed
-
- Burke RE. The use of state-dependent modulation of spinal reflexes as a tool to investigate the organization of spinal interneurons. Exp Brain Res. 1999;128:263–277. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources