

The topoisomerase IIbeta circular clamp arrests transcription and signals a 26S proteasome pathway
- PMID: 12629207
- PMCID: PMC152276
- DOI: 10.1073/pnas.0736401100
The topoisomerase IIbeta circular clamp arrests transcription and signals a 26S proteasome pathway
Abstract
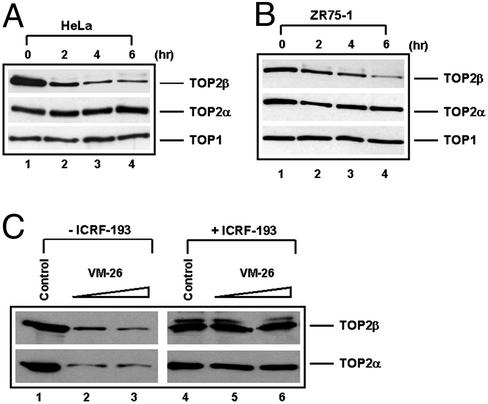
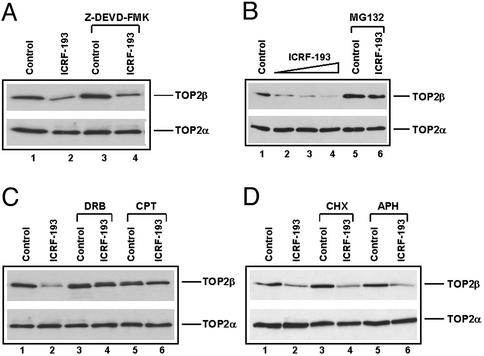
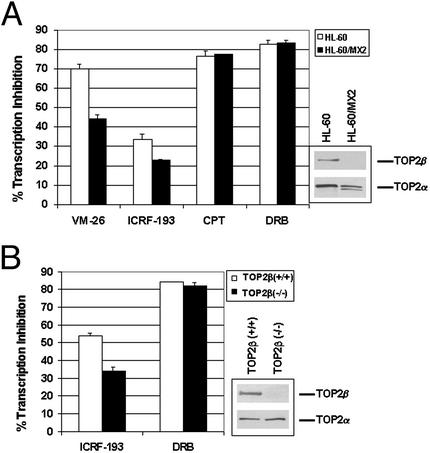
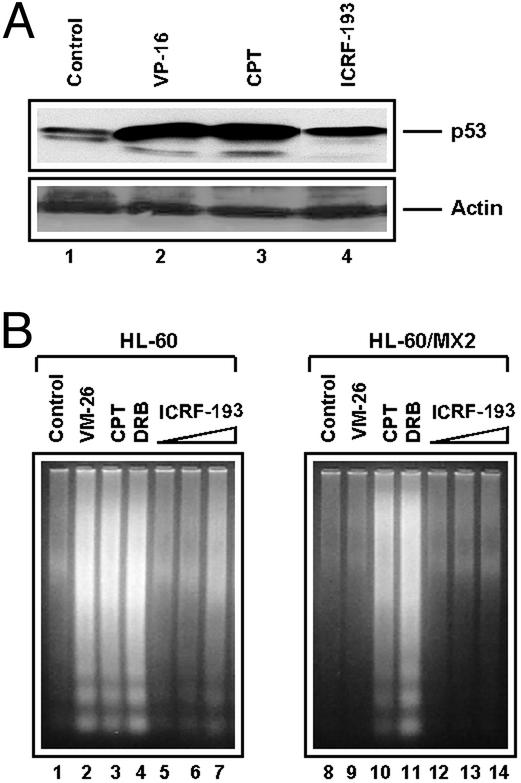
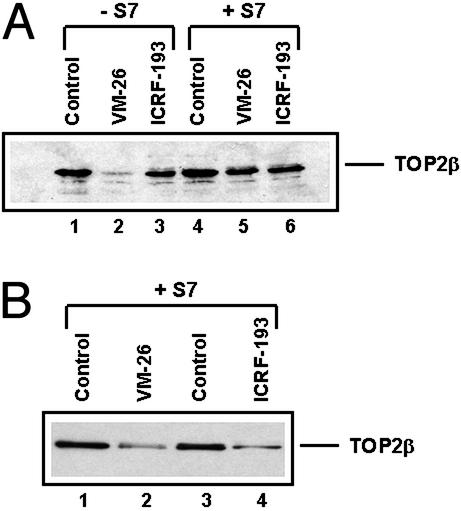
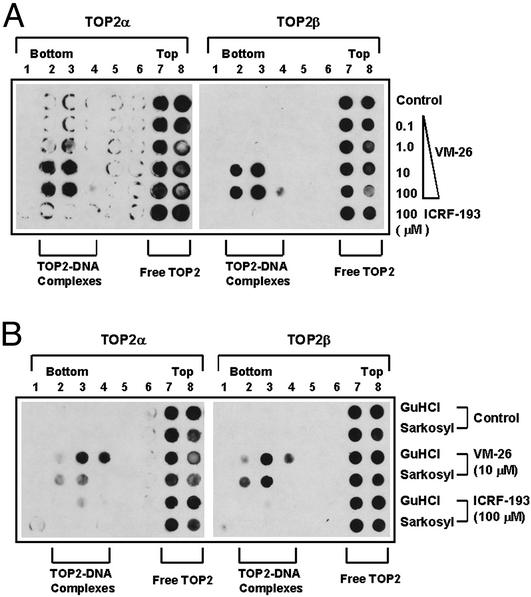
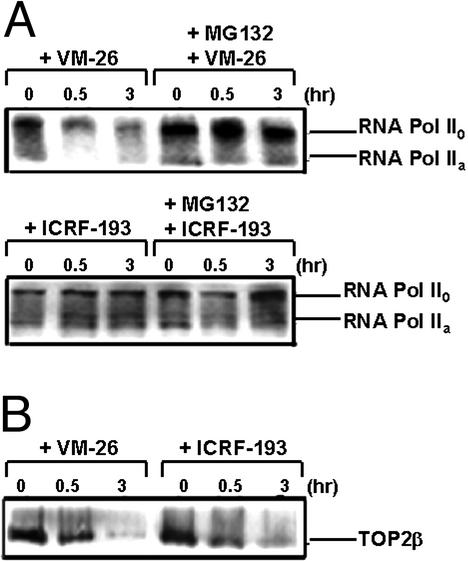
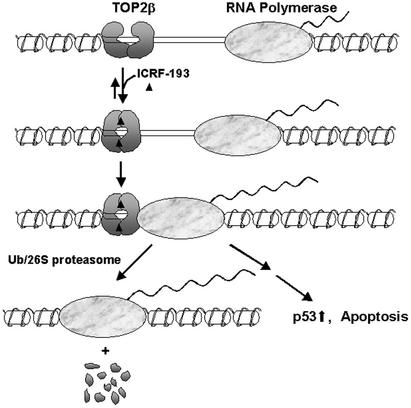
It has been proposed that the topoisomerase II (TOP2)beta-DNA covalent complex arrests transcription and triggers 26S proteasome-mediated degradation of TOP2beta. It is unclear whether the initial trigger for proteasomal degradation is due to DNA damage or transcriptional arrest. In the current study we show that the TOP2 catalytic inhibitor 4,4-(2,3-butanediyl)-bis(2,6-piperazinedione) (ICRF-193), which traps TOP2 into a circular clamp rather than the TOP2-DNA covalent complex, can also arrest transcription. Arrest of transcription, which is TOP2beta-dependent, is accompanied by proteasomal degradation of TOP2beta. Different from TOP2 poisons and other DNA-damaging agents, ICRF-193 did not induce proteasomal degradation of the large subunit of RNA polymerase II. These results suggest that proteasomal degradation of TOP2beta induced by the TOP2-DNA covalent complex or the TOP2 circular clamp is due to transcriptional arrest but not DNA damage. By contrast, degradation of the large subunit of RNA polymerase II is due to a DNA-damage signal.
Figures
References
-
- D'Arpa P, Liu L F. Biochim Biophys Acta. 1989;989:163–177. - PubMed
-
- Li T K, Liu L F. Annu Rev Pharmacol Toxicol. 2001;41:53–77. - PubMed
-
- Kaufmann S H. Biochim Biophys Acta. 1998;1400:195–211. - PubMed
-
- Mao Y, Desai S D, Ting C Y, Hwang J, Liu L F. J Biol Chem. 2001;276:40652–40658. - PubMed
-
- Desai S D, Li T K, Rodriguez-Bauman A, Rubin E H, Liu L F. Cancer Res. 2001;61:5926–5932. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
