

Sequential entry of components of the gene expression machinery into daughter nuclei
- PMID: 12631722
- PMCID: PMC151578
- DOI: 10.1091/mbc.e02-10-0669
Sequential entry of components of the gene expression machinery into daughter nuclei
Erratum in
- Mol Biol Cell. 2003 Apr;14(4):following 1743
Abstract
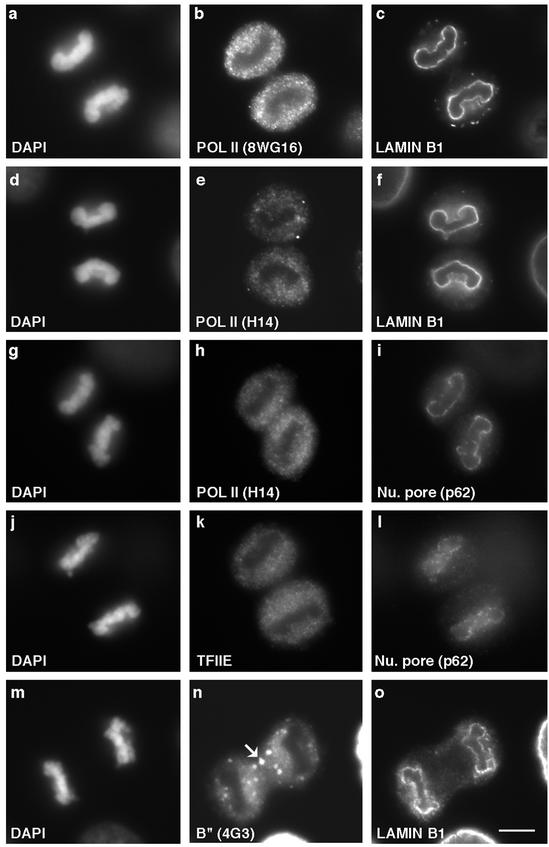
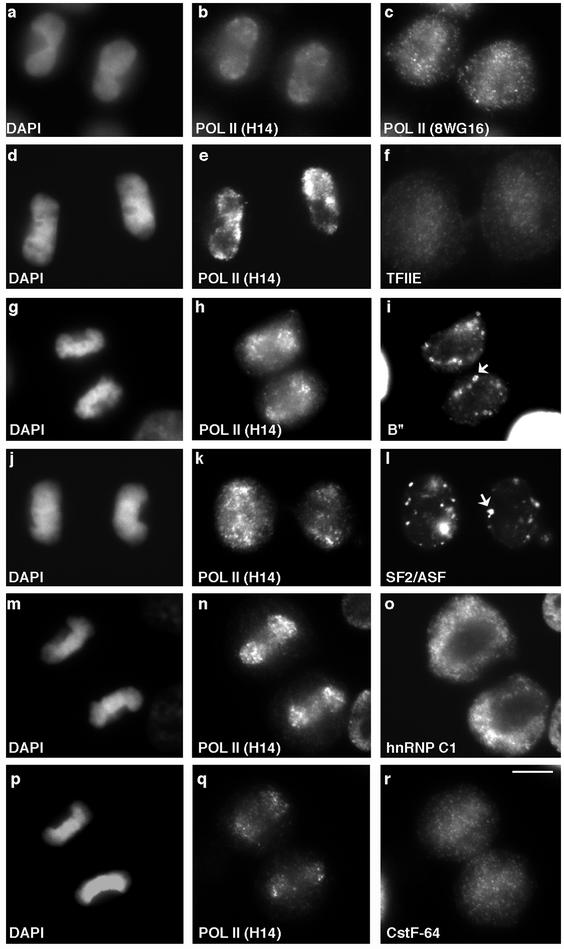
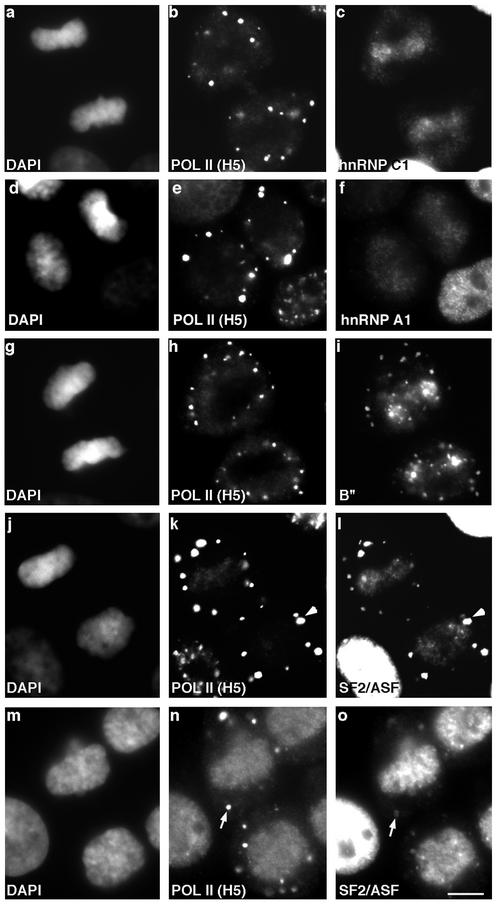
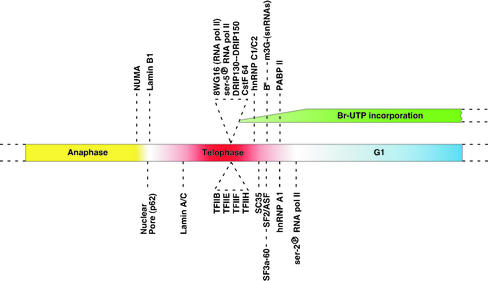
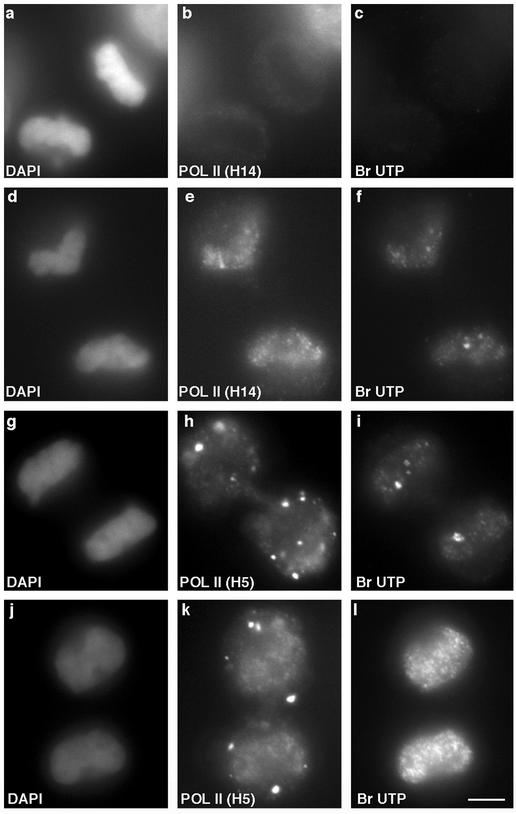
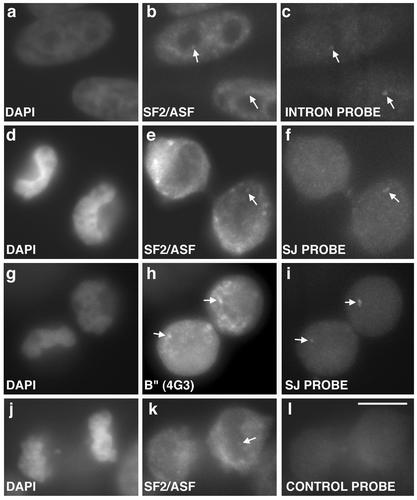
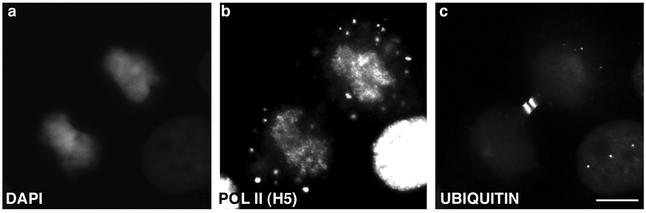
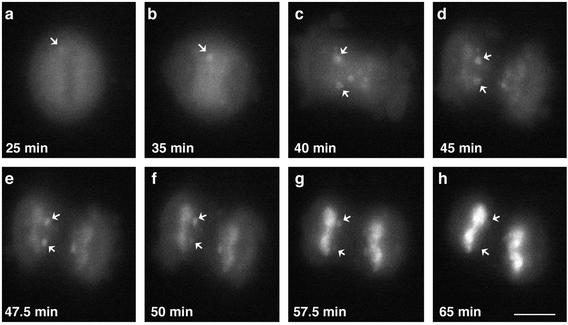
In eukaryotic cells, RNA polymerase II (RNA pol II) transcription and pre-mRNA processing are coordinated events. We have addressed how individual components of the transcription and pre-mRNA processing machinery are organized during mitosis and subsequently recruited into the newly formed daughter nuclei. Interestingly, localization studies of numerous RNA pol II transcription and pre-mRNA processing factors revealed a nonrandom and sequential entry of these factors into daughter nuclei after nuclear envelope/lamina formation. The initiation competent form of RNA pol II and general transcription factors appeared in the daughter nuclei simultaneously, but prior to pre-mRNA processing factors, whereas the elongation competent form of RNA pol II was detected even later. The differential entry of these factors rules out the possibility that they are transported as a unitary complex. Telophase nuclei were competent for transcription and pre-mRNA splicing concomitant with the initial entry of the respective factors. In addition, our results revealed a low turnover rate of transcription and pre-mRNA splicing factors during mitosis. We provide evidence to support a model in which the entry of the RNA pol II gene expression machinery into newly forming daughter nuclei is a staged and ordered process.
Figures
Similar articles
-
Coupling RNA polymerase II transcription with pre-mRNA processing.Curr Opin Cell Biol. 1999 Jun;11(3):347-51. doi: 10.1016/S0955-0674(99)80048-9. Curr Opin Cell Biol. 1999. PMID: 10395561 Review.
-
Linking splicing to Pol II transcription stabilizes pre-mRNAs and influences splicing patterns.PLoS Biol. 2006 Jun;4(6):e147. doi: 10.1371/journal.pbio.0040147. Epub 2006 May 2. PLoS Biol. 2006. PMID: 16640457 Free PMC article.
-
On the importance of being co-transcriptional.J Cell Sci. 2002 Oct 15;115(Pt 20):3865-71. doi: 10.1242/jcs.00073. J Cell Sci. 2002. PMID: 12244124 Review.
-
Coupling of RNA Polymerase II Transcription Elongation with Pre-mRNA Splicing.J Mol Biol. 2016 Jun 19;428(12):2623-2635. doi: 10.1016/j.jmb.2016.04.017. Epub 2016 Apr 20. J Mol Biol. 2016. PMID: 27107644 Free PMC article. Review.
-
Assembly of the nuclear transcription and processing machinery: Cajal bodies (coiled bodies) and transcriptosomes.Mol Biol Cell. 1999 Dec;10(12):4385-402. doi: 10.1091/mbc.10.12.4385. Mol Biol Cell. 1999. PMID: 10588665 Free PMC article.
Cited by
-
Bookmarking by specific and nonspecific binding of FoxA1 pioneer factor to mitotic chromosomes.Genes Dev. 2013 Feb 1;27(3):251-60. doi: 10.1101/gad.206458.112. Epub 2013 Jan 25. Genes Dev. 2013. PMID: 23355396 Free PMC article.
-
Cytosolic condensates rich in polyserine define subcellular sites of tau aggregation.Proc Natl Acad Sci U S A. 2023 Jan 17;120(3):e2217759120. doi: 10.1073/pnas.2217759120. Epub 2023 Jan 10. Proc Natl Acad Sci U S A. 2023. PMID: 36626563 Free PMC article.
-
Bookmarking target genes in mitosis: a shared epigenetic trait of phenotypic transcription factors and oncogenes?Cancer Res. 2014 Jan 15;74(2):420-5. doi: 10.1158/0008-5472.CAN-13-2837. Epub 2014 Jan 9. Cancer Res. 2014. PMID: 24408924 Free PMC article. Review.
-
Differential dynamics of splicing factor SC35 during the cell cycle.J Biosci. 2008 Sep;33(3):345-54. doi: 10.1007/s12038-008-0054-3. J Biosci. 2008. PMID: 19005234
-
Interplay between the alpharetroviral Gag protein and SR proteins SF2 and SC35 in the nucleus.Front Microbiol. 2015 Sep 8;6:925. doi: 10.3389/fmicb.2015.00925. eCollection 2015. Front Microbiol. 2015. PMID: 26441864 Free PMC article.
References
-
- Adam SA. Transport pathways of macromolecules between the nucleus and the cytoplasm. Curr Opin Cell Biol. 1999;11:402–406. - PubMed
-
- Bauren G, Wieslander L. Splicing of Balbiani ring 1 gene pre-mRNA occurs simultaneously with transcription. Cell. 1994;76:183–192. - PubMed
-
- Bensaude O, Bonnet F, Casse C, Dubois MF, Nguyen VT, Palancade B. Regulated phosphorylation of the RNA polymerase II C-terminal domain (CTD) Biochem Cell Biol. 1999;77:249–255. - PubMed
-
- Beyer AL, Osheim YN. Splice site selection, rate of splicing, and alternative splicing on nascent transcripts. Genes Dev. 1988;2:754–765. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
