

In vivo derivation of glucose-competent pancreatic endocrine cells from bone marrow without evidence of cell fusion
- PMID: 12639990
- PMCID: PMC153767
- DOI: 10.1172/JCI16502
In vivo derivation of glucose-competent pancreatic endocrine cells from bone marrow without evidence of cell fusion
Abstract
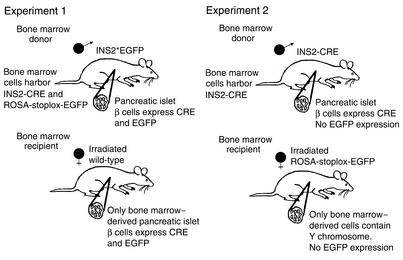
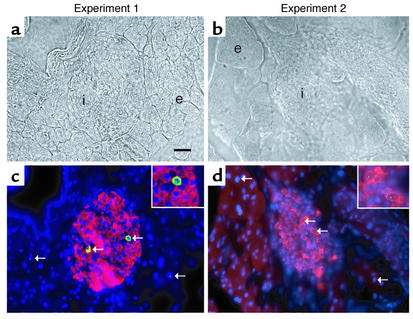
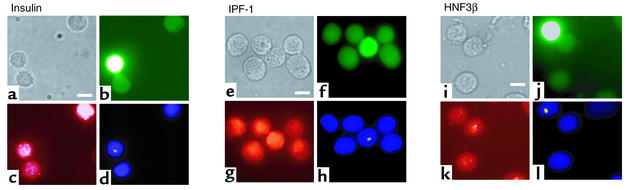
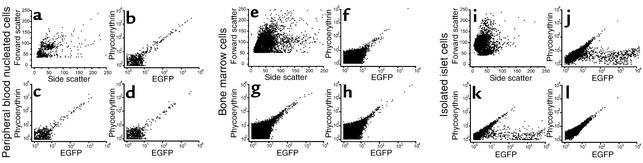
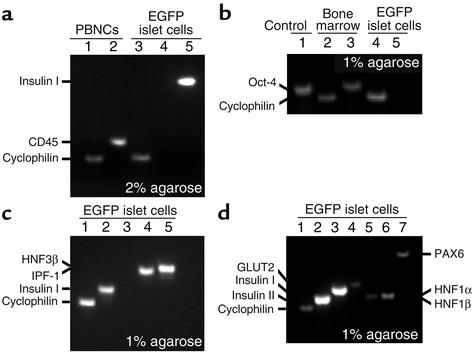
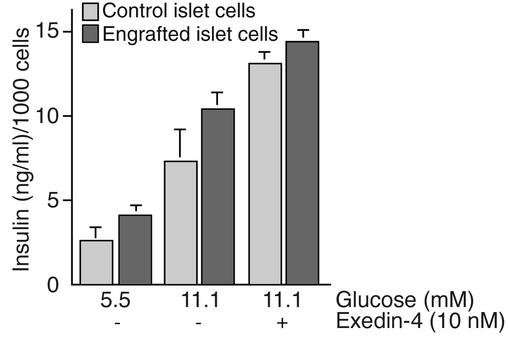
Bone marrow harbors cells that have the capacity to differentiate into cells of nonhematopoietic tissues of neuronal, endothelial, epithelial, and muscular phenotype. Here we demonstrate that bone marrow-derived cells populate pancreatic islets of Langerhans. Bone marrow cells from male mice that express, using a CRE-LoxP system, an enhanced green fluorescent protein (EGFP) if the insulin gene is actively transcribed were transplanted into lethally irradiated recipient female mice. Four to six weeks after transplantation, recipient mice revealed Y chromosome and EGFP double-positive cells in their pancreatic islets. Neither bone marrow cells nor circulating peripheral blood nucleated cells of donor or recipient mice had any detectable EGFP. EGFP-positive cells purified from islets express insulin, glucose transporter 2 (GLUT2), and transcription factors typically found in pancreatic beta cells. Furthermore, in vitro these bone marrow-derived cells exhibit - as do pancreatic beta cells - glucose-dependent and incretin-enhanced insulin secretion. These results indicate that bone marrow harbors cells that have the capacity to differentiate into functionally competent pancreatic endocrine beta cells and that represent a source for cell-based treatment of diabetes mellitus. The results generated with the CRE-LoxP system also suggest that in vivo cell fusion is an unlikely explanation for the "transdifferentiation" of bone marrow-derived cells into differentiated cell phenotypes.
Figures
Comment in
-
Bone marrow: an extra-pancreatic hideout for the elusive pancreatic stem cell?J Clin Invest. 2003 Mar;111(6):799-801. doi: 10.1172/JCI17063. J Clin Invest. 2003. PMID: 12639984 Free PMC article. No abstract available.
References
-
- Rosenberg L, Vinik AI, Pittenger GL, Rafaeloff R, Duguid WP. Islet-cell regeneration in the diabetic hamster pancreas with restoration of normoglycaemia can be induced by a local growth factor(s) Diabetologia. 1996;39:256–262. - PubMed
-
- Rafaeloff R, Barlow SW, Rosenberg L, Vinik AI. Expression of Reg gene in the Syrian golden hamster pancreatic islet regeneration model. Diabetologia. 1995;38:906–913. - PubMed
-
- Stoffers DA, et al. Insulinotropic glucagon-like peptide 1 agonists stimulate expression of homeodomain protein IDX-1 and increase islet size in mouse pancreas. Diabetes. 2000;49:741–748. - PubMed
-
- Guz Y, Nasir I, Teitelman G. Regeneration of pancreatic beta cells from intra-islet precursor cells in an experimental model of diabetes. Endocrinology. 2001;142:4956–4968. - PubMed
-
- Fernandes A, et al. Differentiation of new insulin-producing cells is induced by injury in adult pancreatic islets. Endocrinology. 1997;138:1750–1762. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
