

Review
doi: 10.1083/jcb.200301035.
Polarized growth and organelle segregation in yeast: the tracks, motors, and receptors
Affiliations
- PMID: 12642608
- PMCID: PMC2173777
- DOI: 10.1083/jcb.200301035
Item in Clipboard
Review
Polarized growth and organelle segregation in yeast: the tracks, motors, and receptors
J Cell Biol.
.
Abstract
In yeast, growth and organelle segregation requires formin-dependent assembly of polarized actin cables. These tracks are used by myosin Vs to deliver secretory vesicles for cell growth, organelles for their segregation, and mRNA for fate determination. Several specific receptors have been identified that interact with the cargo-binding tails of the myosin Vs. A recent study implicates specific degradation in the bud of the vacuolar receptor, Vac17, as a mechanism for cell cycle-regulated segregation of this organelle.
Figures
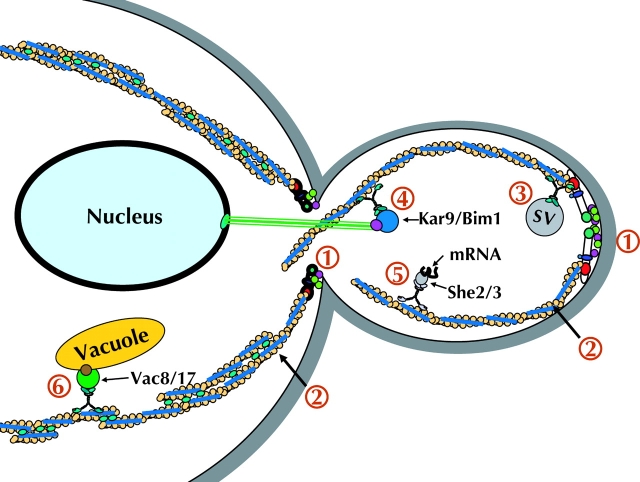
Diagram summarizing the myosin V–based mechanisms of organelle transport. (1) Formins are localized and activated (Bni1 at the bud tip and Bnr1 at the bud neck), which drive the assembly of polarized actin cables. (2) Actin cables are stabilized by tropomyosin (blue) and cross-linking proteins (e.g., fimbrin, green). (3) Myo2 transports post-Golgi secretory vesicles into the bud. (4) Nuclear orientation involves Myo2-dependent transport of Kar9/Bim1 into the bud. (5) Specific mRNAs are selected by the She2/3 complex and transported into the bud by Myo4. (6) Vacuolar elements are moved by Myo2 into the bud through an interaction with Vac8/17. After transport into the bud, Vac17 is degraded (not depicted).
References
-
- Alberts, A.S. 2001. Identification of a carboxyl-terminal diaphanous-related formin homology protein autoregulatory domain. J. Biol. Chem. 276:2824–2830. - PubMed
-
- Beach, D.L., J. Thibodeaux, P. Maddox, E. Yeh, and K. Bloom. 2000. The role of the proteins Kar9 and Myo2 in orienting the mitotic spindle of budding yeast. Curr. Biol. 10:1497–1506. - PubMed
-
- Bobola, N., R.P. Jansen, T.H. Shin, and K. Nasmyth. 1996. Asymmetric accumulation of Ash1p in postanaphase nuclei depends on a myosin and restricts yeast mating-type switching to mother cells. Cell. 84:699–709. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
