

Identification of a new gene essential for germination of Bacillus subtilis spores with Ca2+-dipicolinate
- PMID: 12644503
- PMCID: PMC151495
- DOI: 10.1128/JB.185.7.2315-2329.2003
Identification of a new gene essential for germination of Bacillus subtilis spores with Ca2+-dipicolinate
Abstract
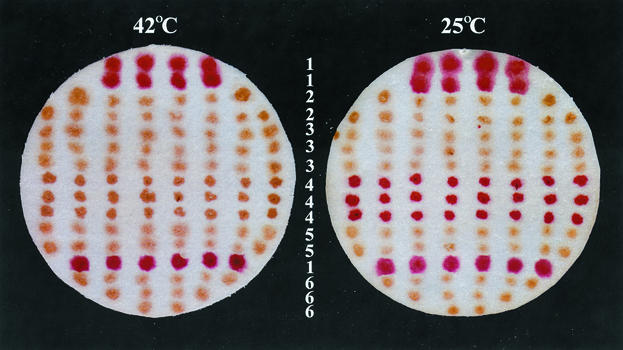
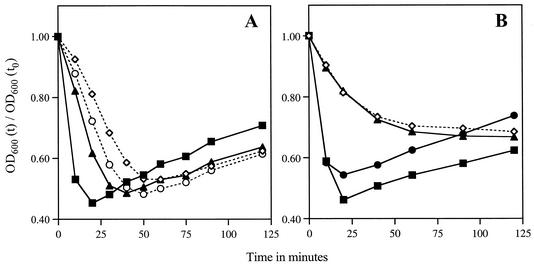
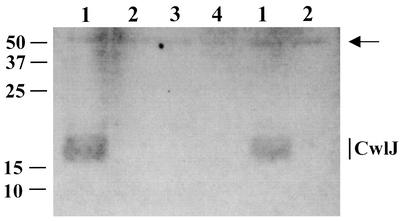
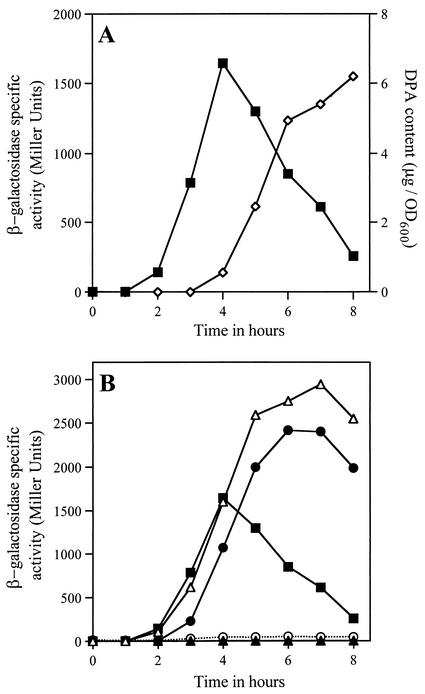
Bacillus subtilis spores can germinate with a 1:1 chelate of Ca(2+) and dipicolinic acid (DPA), a compound present at high levels in the spore core. Using a genetic screen to identify genes encoding proteins that are specifically involved in spore germination by Ca(2+)-DPA, three mutations were identified. One was in the gene encoding the cortex lytic enzyme, CwlJ, that was previously shown to be essential for spore germination by Ca(2+)-DPA. The other two were mapped to an open reading frame, ywdL, encoding a protein of unknown function. Analysis of ywdL expression showed that the gene is expressed during sporulation in the mother cell compartment of the sporulating cell and that its transcription is sigma(E) dependent. Functional characterization of YwdL demonstrated that it is a new spore coat protein that is essential for the presence of CwlJ in the spore coat. Assembly of YwdL itself into the spore coat is dependent on the coat morphogenetic proteins CotE and SpoIVA. However, other than lacking CwlJ, ywdL spores have no obvious defect in their spore coat. Because of the role for YwdL in a part of the spore germination process, we propose renaming ywdL as a spore germination gene, gerQ.
Figures


References
-
- Bagyan, I., L. Casillas-Martinez, and P. Setlow. 1998. The katX gene, which codes for the catalase in spores of Bacillus subtilis, is a forespore-specific gene controlled by σF, and KatX is essential for hydrogen peroxide resistance of the germinating spore. J. Bacteriol. 180:2057-2062. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
