

The Sym35 gene required for root nodule development in pea is an ortholog of Nin from Lotus japonicus
- PMID: 12644653
- PMCID: PMC166866
- DOI: 10.1104/pp.102.016071
The Sym35 gene required for root nodule development in pea is an ortholog of Nin from Lotus japonicus
Abstract
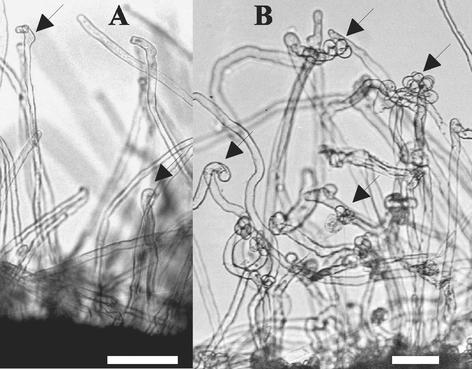
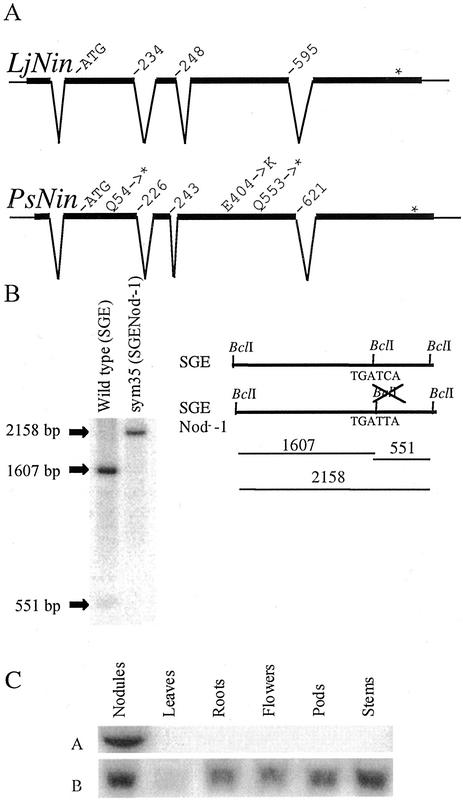
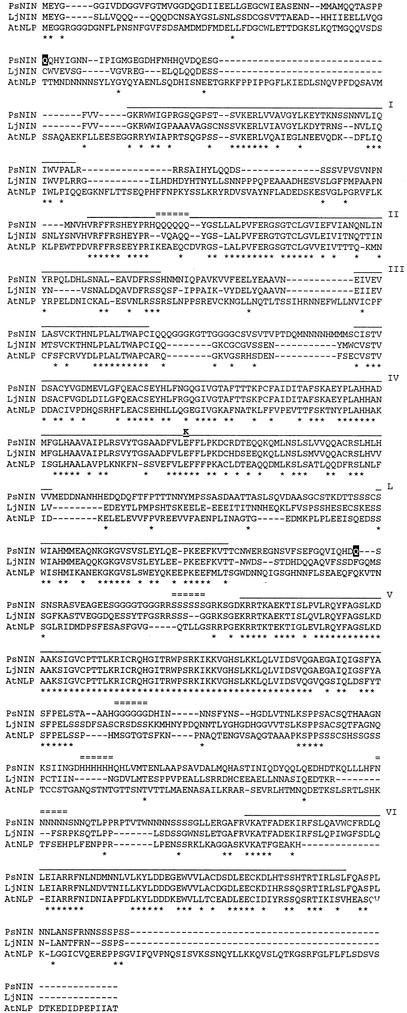
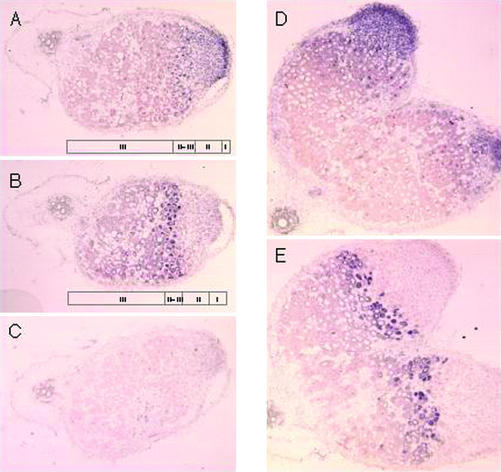
Comparative phenotypic analysis of pea (Pisum sativum) sym35 mutants and Lotus japonicus nin mutants suggested a similar function for the PsSym35 and LjNin genes in early stages of root nodule formation. Both the pea and L. japonicus mutants are non-nodulating but normal in their arbuscular mycorrhizal association. Both are characterized by excessive root hair curling in response to the bacterial microsymbiont, lack of infection thread initiation, and absence of cortical cell divisions. To investigate the molecular basis for the similarity, we cloned and sequenced the PsNin gene, taking advantage of sequence information from the previously cloned LjNin gene. An RFLP analysis on recombinant inbred lines mapped PsNin to the same chromosome arm as the PsSym35 locus and direct evidence demonstrating that PsNin is the PsSym35 gene was subsequently obtained by cosegregation analysis and sequencing of three independent Pssym35 mutant alleles. L. japonicus and pea root nodules develop through different organogenic pathways, so it was of interest to compare the expression of the two orthologous genes during nodule formation. Overall, a similar developmental regulation of the PsNin and LjNin genes was shown by the transcriptional activation in root nodules of L. japonicus and pea. In the indeterminate pea nodules, PsNin is highly expressed in the meristematic cells of zone I and in the cells of infection zone II, corroborating expression of LjNin in determinate nodule primordia. At the protein level, seven domains, including the putative DNA binding/dimerization RWP-RK motif and the PB1 heterodimerization domain, are conserved between the LjNIN and PsNIN proteins.
Figures

References
-
- Bladergroen MR, Spaink HP. Genes and signal molecules involved in the Rhizobia-Leguminosae symbiosis. Curr Opin Plant Biol. 1998;1:353–359. - PubMed
-
- Bonfante P, Genre A, Faccio A, Martini I, Schauser L, Stougaard J, Webb J, Parniske M. The Lotus japonicus LjSym4 gene is required for the successful symbiotic infection of root epidermal cells. Mol Plant-Microbe Interact. 2000;13:1109–1120. - PubMed
-
- Borisov AY, Barmicheva EM, Jacobi LM, Tsyganov VE, Voroshilova VA, Tikhonovich IA. Pea (Pisum sativum L.) mendelian genes controlling development of nitrogen-fixing nodules and arbuscular mycorrhiza. Czech J Genet Plant Breed. 2000;36:106–110.
-
- Borisov AY, Rozov SM, Tsyganov VE, Morzhina EV, Lebsky VK, Tikhonovich IA. Sequential functioning of Sym-13 and Sym-31, two genes affecting symbisome development in root nodules of pea (Pisum sativum L.) Mol Gen Genet. 1997;254:592–598. - PubMed
-
- Downie JA, Walker SA. Plant responses to nodulation factors. Curr Opin Plant Biol. 1999;2:483–489. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
