

Isolation and properties of floral defensins from ornamental tobacco and petunia
- PMID: 12644678
- PMCID: PMC166888
- DOI: 10.1104/pp.102.016626
Isolation and properties of floral defensins from ornamental tobacco and petunia
Abstract
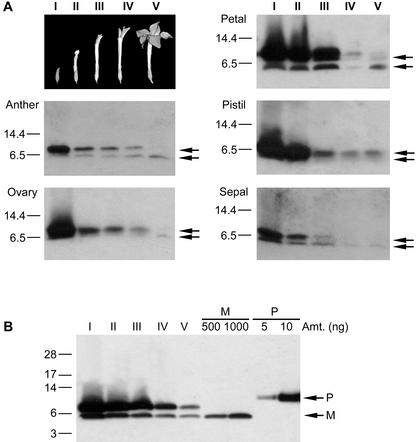
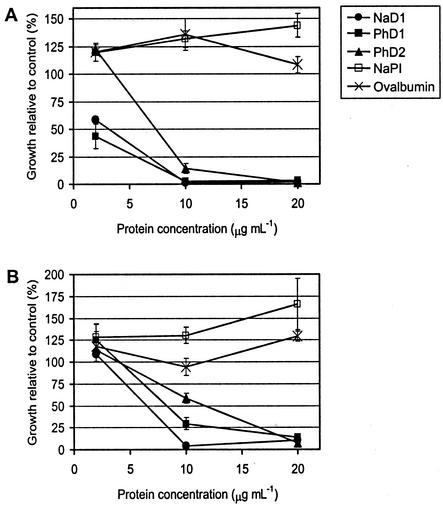
The flowers of the solanaceous plants ornamental tobacco (Nicotiana alata) and petunia (Petunia hybrida) produce high levels of defensins during the early stages of development. In contrast to the well-described seed defensins, these floral defensins are produced as precursors with C-terminal prodomains of 27 to 33 amino acids in addition to a typical secretion signal peptide and central defensin domain of 47 or 49 amino acids. Defensins isolated from N. alata and petunia flowers lack the C-terminal domain, suggesting that it is removed during or after transit through the secretory pathway. Immunogold electron microscopy has been used to demonstrate that the N. alata defensin is deposited in the vacuole. In addition to the eight canonical cysteine residues that define the plant defensin family, the two petunia defensins have an extra pair of cysteines that form a fifth disulfide bond and hence define a new subclass of this family of proteins. Expression of the N. alata defensin NaD1 is predominantly flower specific and is most active during the early stages of flower development. NaD1 transcripts accumulate in the outermost cell layers of petals, sepals, anthers, and styles, consistent with a role in protection of the reproductive organs against potential pathogens. The floral defensins inhibit the growth of Botrytis cinerea and Fusarium oxysporum in vitro, providing further support for a role in protection of floral tissues against pathogen invasion.
Figures




Similar articles
-
Inhibition of cereal rust fungi by both class I and II defensins derived from the flowers of Nicotiana alata.Mol Plant Pathol. 2014 Jan;15(1):67-79. doi: 10.1111/mpp.12066. Epub 2013 Sep 10. Mol Plant Pathol. 2014. PMID: 24015961 Free PMC article.
-
Snakin-1, a peptide from potato that is active against plant pathogens.Mol Plant Microbe Interact. 1999 Jan;12(1):16-23. doi: 10.1094/MPMI.1999.12.1.16. Mol Plant Microbe Interact. 1999. PMID: 9885189
-
Downregulation of the Petunia hybrida alpha-expansin gene PhEXP1 reduces the amount of crystalline cellulose in cell walls and leads to phenotypic changes in petal limbs.Plant Cell. 2004 Feb;16(2):295-308. doi: 10.1105/tpc.018705. Epub 2004 Jan 23. Plant Cell. 2004. PMID: 14742876 Free PMC article.
-
The evolution, function and mechanisms of action for plant defensins.Semin Cell Dev Biol. 2019 Apr;88:107-118. doi: 10.1016/j.semcdb.2018.02.004. Epub 2018 Feb 23. Semin Cell Dev Biol. 2019. PMID: 29432955 Review.
-
Unraveling the regulation of floral fragrance biosynthesis.Plant Signal Behav. 2011 Mar;6(3):378-81. doi: 10.4161/psb.6.3.14339. Plant Signal Behav. 2011. PMID: 21673507 Free PMC article. Review.
Cited by
-
Hyperpolarisation of Mitochondrial Membranes Is a Critical Component of the Antifungal Mechanism of the Plant Defensin, Ppdef1.J Fungi (Basel). 2024 Jan 7;10(1):54. doi: 10.3390/jof10010054. J Fungi (Basel). 2024. PMID: 38248963 Free PMC article.
-
Phosphoinositide-mediated oligomerization of a defensin induces cell lysis.Elife. 2014 Apr 1;3:e01808. doi: 10.7554/eLife.01808. Elife. 2014. PMID: 24692446 Free PMC article.
-
Antimicrobial Peptides from Plants.Pharmaceuticals (Basel). 2015 Nov 16;8(4):711-57. doi: 10.3390/ph8040711. Pharmaceuticals (Basel). 2015. PMID: 26580629 Free PMC article. Review.
-
Mechanisms of Antimicrobial Peptides from Bagasse against Human Pathogenic Bacteria.Antibiotics (Basel). 2023 Feb 23;12(3):448. doi: 10.3390/antibiotics12030448. Antibiotics (Basel). 2023. PMID: 36978315 Free PMC article.
-
Ha-DEF1, a sunflower defensin, induces cell death in Orobanche parasitic plants.Planta. 2007 Aug;226(3):591-600. doi: 10.1007/s00425-007-0507-1. Epub 2007 Mar 21. Planta. 2007. PMID: 17375322
References
-
- Anderson MA, Harris PJ, Bönig I, Clarke AE. Immunogold localization of α-l-arabinofuranosyl residues in pollen tubes of Nicotiana alata Link et Otto. Planta. 1987;171:438–442. - PubMed
-
- Bloch C, Jr, Richardson M. A new family of small (5 kD) protein inhibitors of insect alpha-amylases from seeds or sorghum (Sorghum bicolor Moench) have sequence homologies with wheat γ-purothionins. FEBS Lett. 1991;279:101–104. - PubMed
-
- Bohlmann H. The role of thionins in plant protection. Crit Rev Plant Sci. 1994;13:1–16.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
