

The role of biotin in regulating 3-methylcrotonyl-coenzyme a carboxylase expression in Arabidopsis
- PMID: 12644697
- PMCID: PMC166907
- DOI: 10.1104/pp.013243
The role of biotin in regulating 3-methylcrotonyl-coenzyme a carboxylase expression in Arabidopsis
Abstract
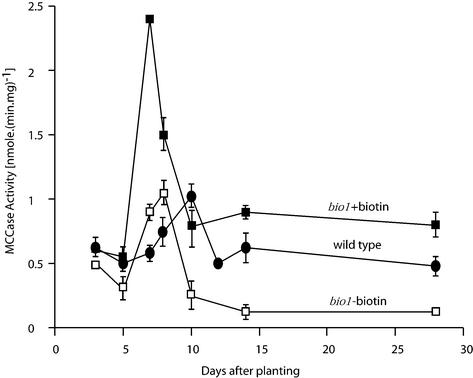
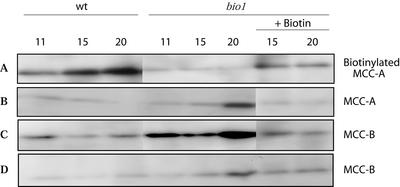
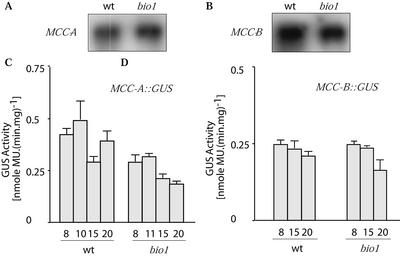
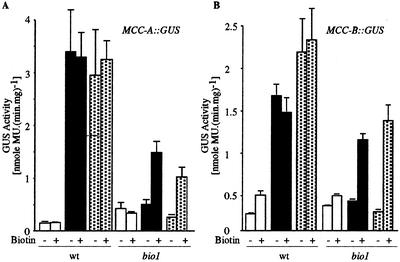
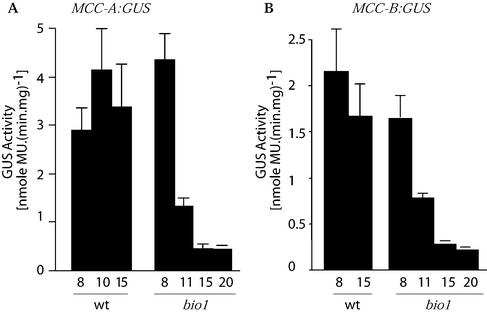
As a catalytic cofactor, biotin has a critical role in the enzymological mechanism of a number of enzymes that are essential in both catabolic and anabolic metabolic processes. In this study we demonstrate that biotin has additional non-catalytic functions in regulating gene expression in plants, which are biotin autotrophic organisms. Biotin controls expression of the biotin-containing enzyme, methylcrotonyl-coenzyme A (CoA) carboxylase by modulating the transcriptional, translational and/or posttranslational regulation of the expression of this enzyme. The bio1 mutant of Arabidopsis, which is blocked in the de novo biosynthesis of biotin, was used to experimentally alter the biotin status of this organism. In response to the bio1-associated depletion of biotin, the normally biotinylated A-subunit of methylcrotonyl-CoA carboxylase (MCCase) accumulates in its inactive apo-form, and both MCCase subunits hyperaccumulate. This hyperaccumulation occurs because the translation of each subunit mRNA is enhanced and/or because the each protein subunit becomes more stable. In addition, biotin affects the accumulation of distinct charge isoforms of MCCase. In contrast, in response to metabolic signals arising from the alteration in the carbon status of the organism, biotin modulates the transcription of the MCCase genes. These experiments reveal that in addition to its catalytic role as an enzyme cofactor, biotin has multiple roles in regulating gene expression.
Figures
References
-
- Alban C, Job D, Douce R. Biotin metabolism in plants. Annu Rev Plant Physiol Plant Mol Biol. 2000;51:17–47. - PubMed
-
- Baldet P, Alban C, Axiotis S, Douce R. Biotin biosynthesis in plants: identification of intermediates. Eur J Biochem Arch Biochem Biophys. 1993;303:67–73.
-
- Baldet P, Alban C, Douce R. Biotin synthesis in higher plants: purification and characterization of bioB gene product equivalent from Arabidopsis thaliana overexpressed in Escherichia coli and its subcellular localization in pea leaf cells. FEBS Lett. 1997;419:206–210. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
