

Induction of G1 cycle arrest in T lymphocytes results in increased extracellular levels of beta-chemokines: a strategy to inhibit R5 HIV-1
- PMID: 12644703
- PMCID: PMC153068
- DOI: 10.1073/pnas.0630584100
Induction of G1 cycle arrest in T lymphocytes results in increased extracellular levels of beta-chemokines: a strategy to inhibit R5 HIV-1
Abstract
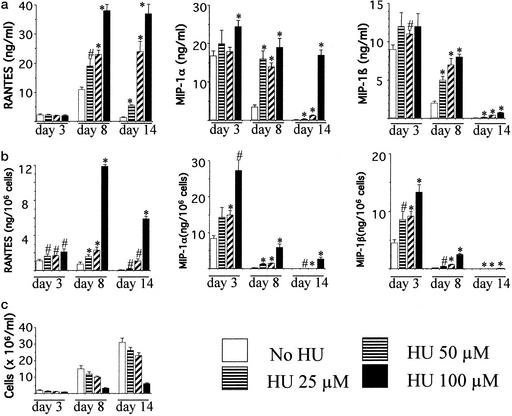
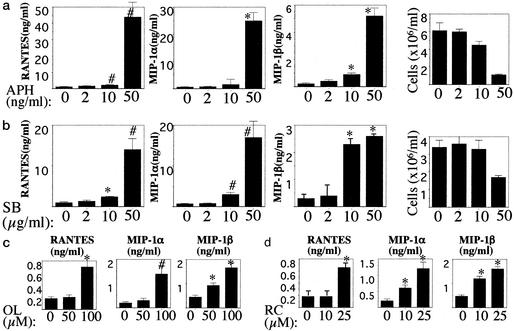
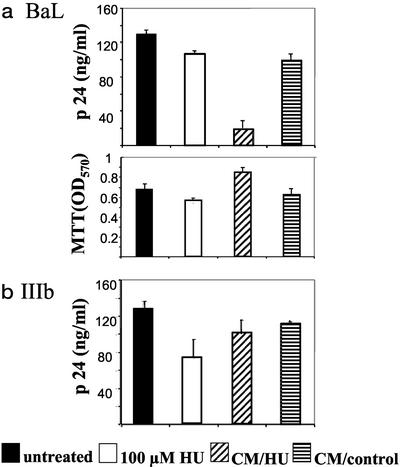
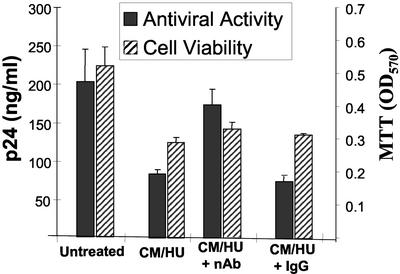
The beta-chemokines RANTES (regulated on activation, normal T cell expressed and secreted), macrophage inflammatory protein-1alpha (MIP-1alpha), and MIP-1beta are the natural ligands of the HIV-1 coreceptor CCR5 and compete with the virus for receptor binding. We show that secretion of the beta-chemokines by activated lymphocytes starts before cellular DNA synthesis is detected and demonstrate that transient prolongation of the G(1) phase of the cell cycle by treatment with cytostatic drugs results in increased levels of the three chemokines in culture supernatants. Supernatants collected from peripheral blood mononuclear cells exposed to hydroxyurea, which arrests the cell cycle in late G(1), contained high levels of beta-chemokines. These supernatants were able to inhibit HIV-1 replication when added to cultures of infected lymphocytes. The observed antiviral effect likely was due to the increased levels of beta-chemokines RANTES, MIP-1alpha, and MIP-1beta because (i) supernatants greatly inhibited the replication of HIV-1 BaL, whereas they affected HIV-1 IIIb replication only slightly; (ii) neutralizing antibodies against the chemokines abrogated the antiviral effect of the supernatants; and (iii) the hydroxyurea concentrations shown to up-regulate chemokine levels were not sufficient to inhibit virus replication by depletion of intracellular nucleotide pools. Although antiviral properties have been reported previously for the cytostatic agents shown here to up-regulate beta-chemokine levels, our results provide an additional mechanism by which these drugs may exert antiviral activity. In summary, increased extracellular levels of anti-HIV-1 beta-chemokines resulting from transient prolongation of the G(1) phase of the lymphocyte cell cycle by treatment with cytostatic drugs may help to control the replication of CCR5-using strains of HIV-1.
Figures


References
-
- Cocchi F, DeVico A L, Garzino-Demo A, Arya S K, Gallo R C, Lusso P. Science. 1995;270:1811–1815. - PubMed
-
- Paxton W A, Martin S R, Tse D, O'Brien J, Skurnick J, Van Devanter N L, Padain N, Braun J F, Kotler S M, Wolinsky S M, et al. Nat Med. 1996;2:412–417. - PubMed
-
- Ferbas J, Giorgi J V, Amini S, Grovit-Ferbas K, Wiley D J, Detels R, Plaeger S. J Infect Dis. 2000;182:1247–1250. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
