

Noncovalent scFv multimers of tumor-targeting anti-Lewis(y) hu3S193 humanized antibody
- PMID: 12649432
- PMCID: PMC2323837
- DOI: 10.1110/ps.0228503
Noncovalent scFv multimers of tumor-targeting anti-Lewis(y) hu3S193 humanized antibody
Abstract
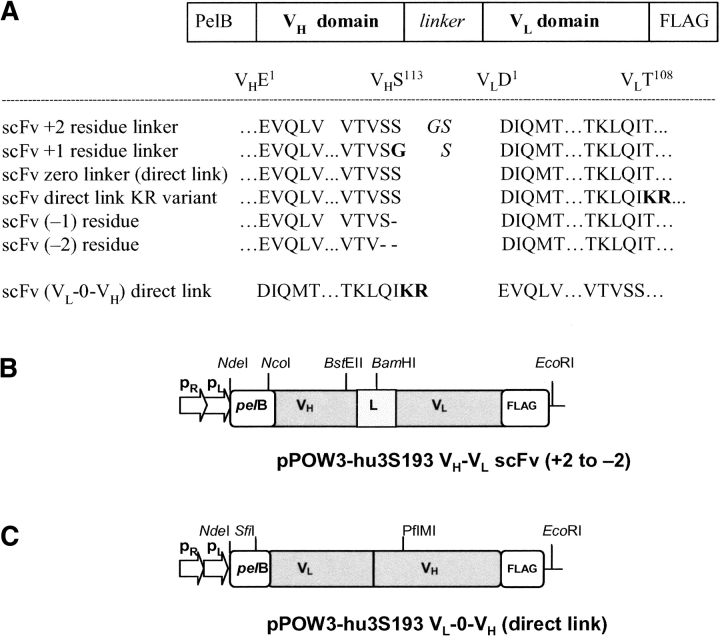
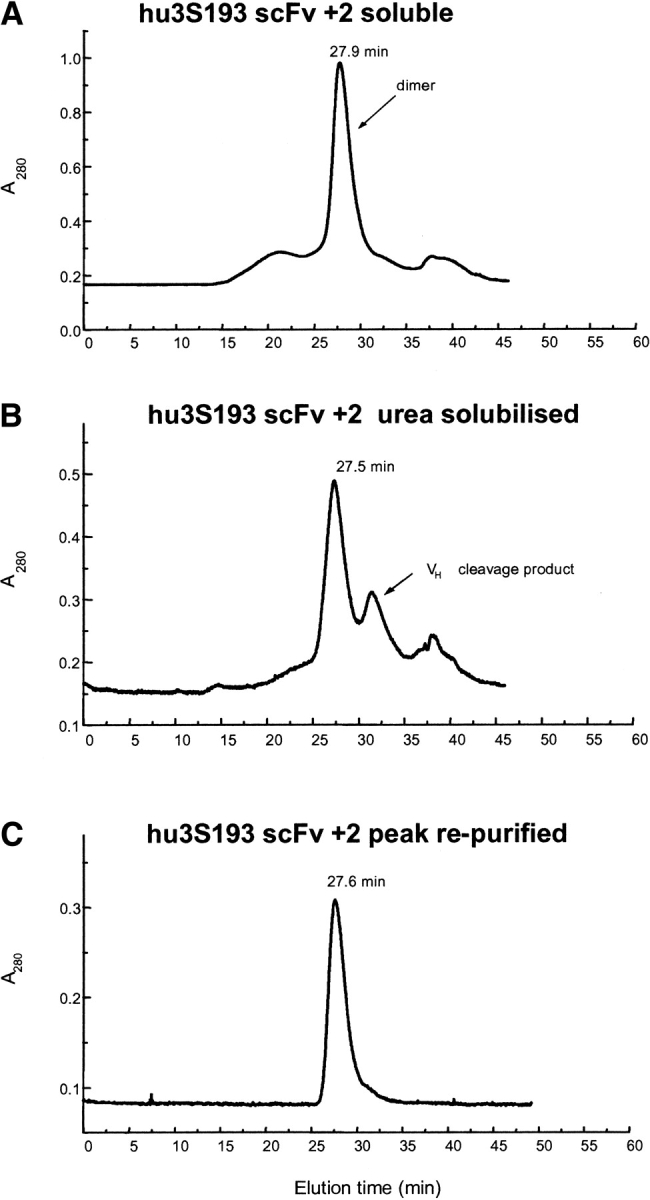
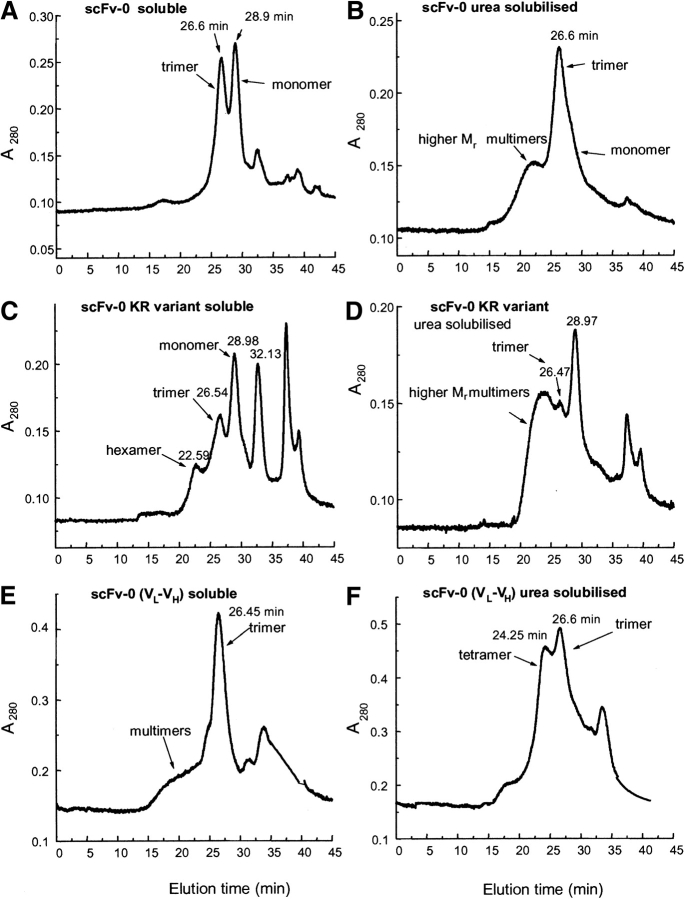
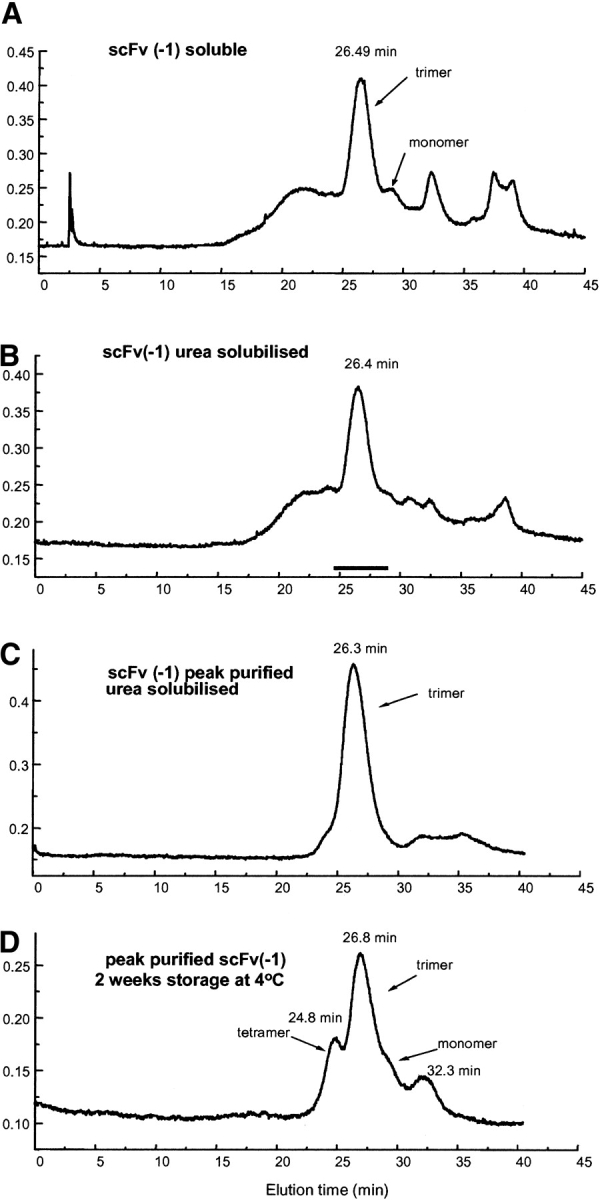
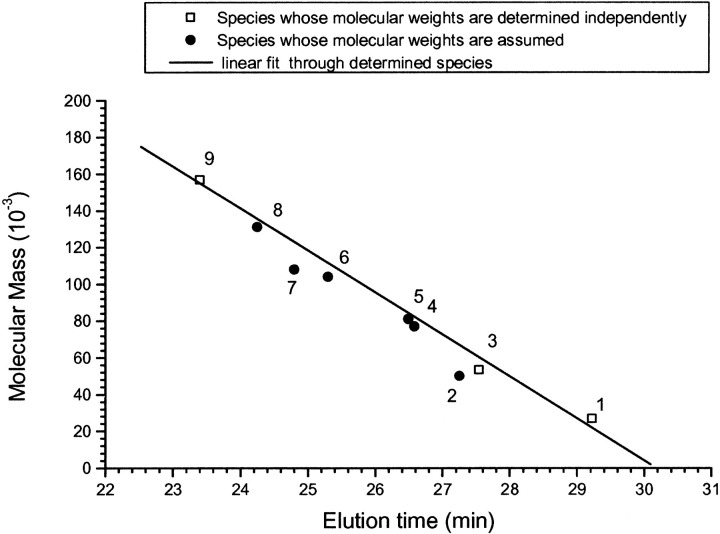
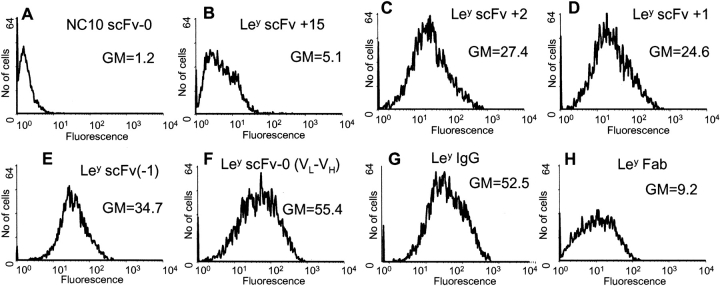
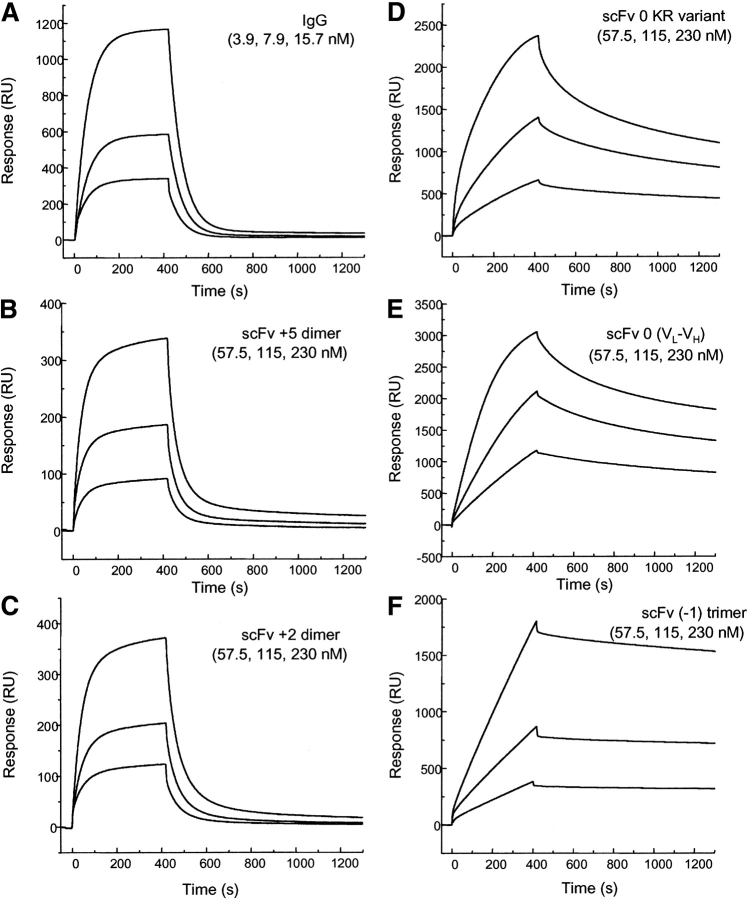
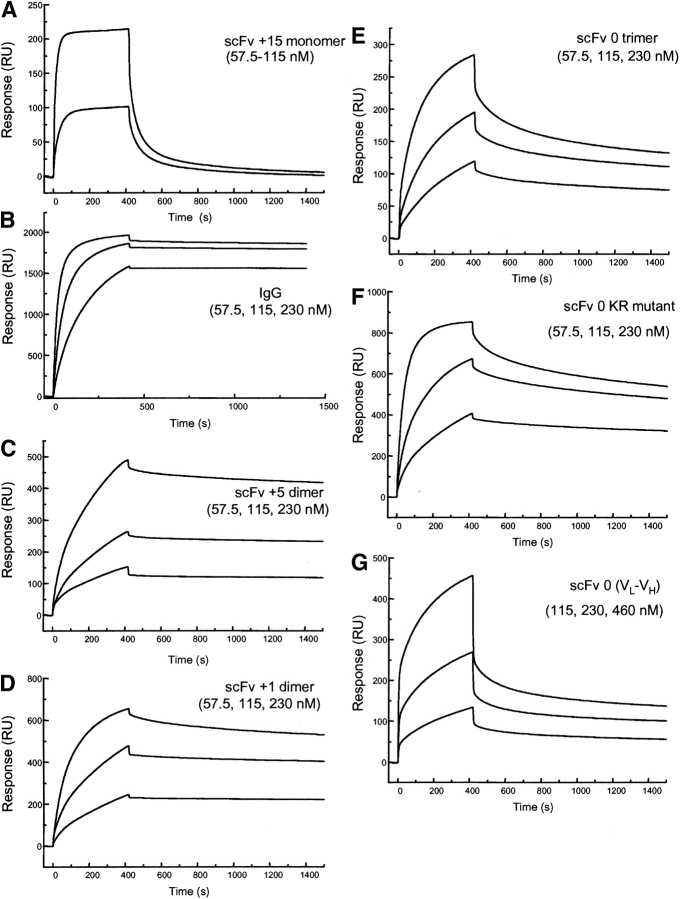
Single-chain variable fragments (scFvs) of anti-Lewis(y) hu3S193 humanized antibody were constructed by joining the V(H) and V(L) domains with either +2 residues, +1 residue, or by directly linking the domains. In addition two constructs were synthesized in which one or two C-terminal residues of the V(H) domain were removed (-1 residue, -2 residue) and then joined directly to the V(L) domain. An scFv construct in the reverse orientation with the V(L) joined directly to the V(H) domain was also synthesized. Upon transformation into Escherichia coli all scFv constructs expressed active protein. Binding activity, multimeric status, and multivalent properties were assessed by flow cytometry, size exclusion chromatography, and biosensor analysis. The results for hu3S193 scFvs are consistent with the paradigm that scFvs with a linker of +3 residues or more associate to form a non-covalent dimer, and those with a shorter linker or directly linked associate predominantly to form a non-covalent trimer and tetramer that are in equilibrium. While the association of V domains to form either a dimer or trimer/tetramer is governed by the length of the linker, the stability of the trimer/tetramer in the equilibrium mixture is dependent on the affinity of the interaction of the individual V domains to associate to form the larger Fv module.
Figures
References
-
- Adams, G.P. and Schier, R. 1999. Generating improved single-chain Fv molecules for tumor targeting. J. Immunol. Methods 231 249–260. - PubMed
-
- Adams, G.P., Schier, R., Marshall, K., Wolf, E.J., McCall, A.M., Marks, J.D., and Weiner, L.M. 1998b. Increased affinity leads to improved selective tumor delivery of single-chain Fv antibodies. Cancer Res. 58 485–490. - PubMed
-
- Adams, G.P., Shaller, C.C., Chappell, L.L., Wu, C., Horak, E.M., Simmons, H.H., Litwin, S., Marks, J.D., Weiner, L.M., and Brechbiel, M.W. 2000. Delivery of the α-emitting radioisotope bismuth-213 to solid tumors via single-chain Fv and diabody molecules. Nucl. Med. Biol. 27 339–346. - PubMed
-
- Atwell, J.L., Breheney, K.A., Lawrence, L.J., McCoy, A.J., Kortt, A.A., and Hudson, P.J. 1999. scFv multimers of the anti-neuraminidase antibody NC10: Length of the linker between VH and VL domains dictates precisely the transition between diabodies and triabodies. Protein Eng. 12 597–604. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources