

Sequence and structural requirements for optimal guide RNA-directed insertional editing within Leishmania tarentolae
- PMID: 12649498
- PMCID: PMC1370413
- DOI: 10.1261/rna.2175703
Sequence and structural requirements for optimal guide RNA-directed insertional editing within Leishmania tarentolae
Abstract
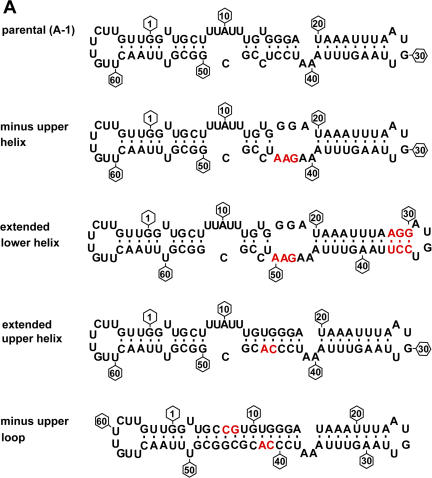
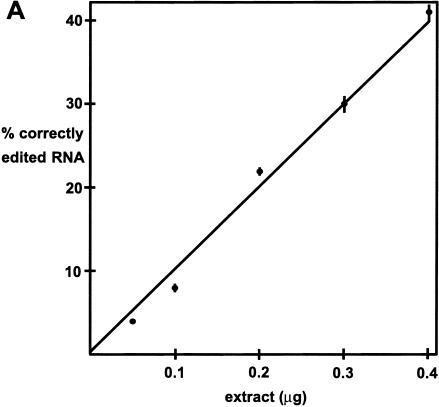
The coding sequence of several mitochondrial mRNAs of the trypanosomatid family of protozoa is created by the guide RNA-directed insertion and deletion of uridylates (Us). Selection-amplification was used to explore the sequence and structure of the guide RNA and mRNA required for efficient insertional editing within a mitochondrial extract prepared from Leishmania tarentolae. This study identifies several novel features of the editing reaction in addition to several that are consistent with the previous mutagenesis and phylogenetic analysis of the reaction in Trypanosoma brucei, a distantly related trypanosomatid. Specifically, there is a strong bias against cytidines 5' of the editing sites and guanosines immediately 3' of guiding nucleotides. U insertions are directed both 5' and 3' of a genomically encoded U, which was previously assumed not to occur. Base pairing immediately flanking an editing site can significantly stimulate the editing reaction and affect the reaction fidelity but is not essential. Likewise, single-stranded RNA in the region upstream of the editing site, not necessarily immediately adjacent, can facilitate editing but is also not essential. The editing of an RNA containing many of the optimal features is linear with increasing quantities of extract permitting specific activity measurements to be made that are not possible with previously described T. brucei and L. tarentolae assays. The reaction catalyzed by the L. tarentolae extract can be highly accurate, which does not support a proposed model for editing that was based largely on the inaccuracy of an earlier in vitro reaction.
Figures


















Similar articles
-
Guide RNA-directed uridine insertion RNA editing in vitro.EMBO J. 1996 Dec 2;15(23):6758-65. EMBO J. 1996. PMID: 8978701 Free PMC article.
-
Direct visualisation of RNA editing within a Leishmania tarentolae mitochondrial extract.Int J Parasitol. 2002 Jun 15;32(7):859-66. doi: 10.1016/s0020-7519(02)00015-2. Int J Parasitol. 2002. PMID: 12062557
-
In vitro uridine insertion RNA editing mediated by cis-acting guide RNAs.RNA. 1999 May;5(5):656-69. doi: 10.1017/s1355838299982250. RNA. 1999. PMID: 10334336 Free PMC article.
-
Kinetoplastid RNA editing: complexes and catalysts.Curr Opin Chem Biol. 1997 Oct;1(3):340-6. doi: 10.1016/s1367-5931(97)80071-3. Curr Opin Chem Biol. 1997. PMID: 9667870 Review.
-
RNA editing in mitochondria of Leishmania tarentolae and Crithidia fasciculata.Semin Cell Biol. 1993 Aug;4(4):241-9. doi: 10.1006/scel.1993.1029. Semin Cell Biol. 1993. PMID: 7694672 Review.
Cited by
-
Identification of specific inhibitors for a trypanosomatid RNA editing reaction.RNA. 2010 Dec;16(12):2435-41. doi: 10.1261/rna.2347310. Epub 2010 Oct 12. RNA. 2010. PMID: 20940340 Free PMC article.
-
Minimal pre-mRNA substrates with natural and converted sites for full-round U insertion and U deletion RNA editing in trypanosomes.Nucleic Acids Res. 2005 Nov 23;33(20):6610-20. doi: 10.1093/nar/gki943. Print 2005. Nucleic Acids Res. 2005. PMID: 16306234 Free PMC article.
-
Complete cycles of bloodstream trypanosome RNA editing in vitro.RNA. 2004 Jun;10(6):914-20. doi: 10.1261/rna.5157704. RNA. 2004. PMID: 15146075 Free PMC article.
-
An electrochemiluminescent aptamer switch for a high-throughput assay of an RNA editing reaction.RNA. 2009 Oct;15(10):1929-38. doi: 10.1261/rna.1720209. Epub 2009 Aug 20. RNA. 2009. PMID: 19696159 Free PMC article.
-
RNA editing complex interactions with a site for full-round U deletion in Trypanosoma brucei.RNA. 2006 Jul;12(7):1219-28. doi: 10.1261/rna.2295706. Epub 2006 May 11. RNA. 2006. PMID: 16690999 Free PMC article.
References
-
- Aphasizhev, R., Sbicego, S., Peris, M., Jang, S.H., Aphasizheva, I., Simpson, A.M., Rivlin, A., and Simpson, L. 2002. Trypanosome mitochondrial 3′ terminal uridylyl transferase (TUTase): The key enzyme in U insertion/deletion RNA editing. Cell 108: 637–648. - PubMed
-
- Benne, R. 1992. RNA editing in trypanosomes. The us(e) of guide RNAs. Mol. Biol. Rep. 16: 217–227. - PubMed
-
- Benne, R., Van den Burg, J., Brakenhoff, J.P., Sloof, P., Van Boom, J.H., and Tromp, M.C. 1986. Major transcript of the frameshifted coxII gene from trypanosome mitochondria contains four nucleotides that are not encoded in the DNA. Cell 46: 819–826. - PubMed
-
- Blum, B., Bakalara, N., and Simpson, L. 1990. A model for RNA editing in kinetoplastid mitochondria: “Guide” RNA molecules transcribed from maxicircle DNA provide the edited information. Cell 60: 189–198. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources