

Essential role for caspase 8 in T-cell homeostasis and T-cell-mediated immunity
- PMID: 12654726
- PMCID: PMC196031
- DOI: 10.1101/gad.1063703
Essential role for caspase 8 in T-cell homeostasis and T-cell-mediated immunity
Abstract
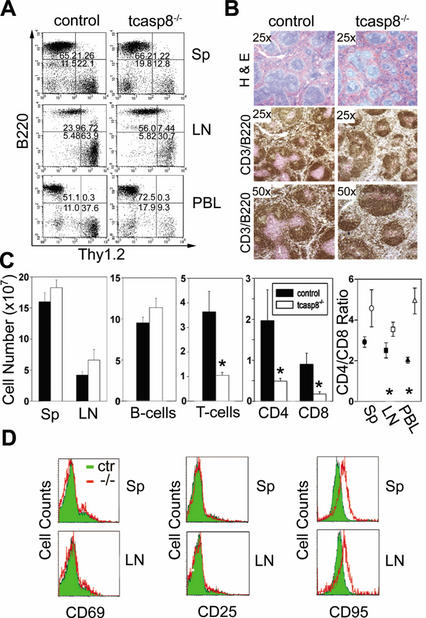
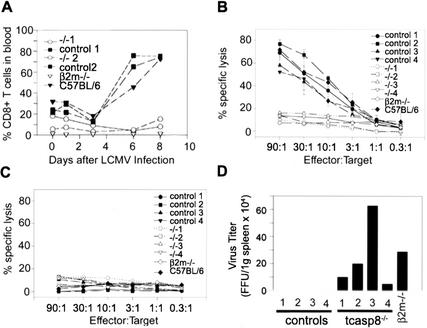
Defects in death receptor-mediated apoptosis have been linked to cancer and autoimmune disease in humans. The in vivo role of caspase 8, a component of this pathway, has eluded analysis in postnatal tissues because of the lack of an appropriate animal model. Targeted disruption of caspase 8 is lethal in utero. We generated mice with a targeted caspase 8 mutation that is restricted to the T-cell lineage. Despite normal thymocyte development in the absence of caspase 8, we observed a marked decrease in the number of peripheral T-cells and impaired T-cell response ex vivo to activation stimuli. caspase 8 ablation protected thymocytes and activated T-cells from CD95 ligand but not anti-CD3-induced apoptosis, or apoptosis activated by agents that are known to act through the mitochondria. caspase 8 mutant mice were unable to mount an immune response to viral infection, indicating that caspase 8 deletion in T-cells leads to immunodeficiency. These findings identify an essential, cell-stage-specific role for caspase 8 in T-cell homeostasis and T-cell-mediated immunity. This is consistent with the recent identification of caspase 8 mutations in human immunodeficiency.
Figures





References
-
- Ashkenazi A. Targeting death and decoy receptors of the tumour-necrosis factor superfamily. Nat Rev Cancer. 2002;2:420–430. - PubMed
-
- Berry DM, Benn SJ, Cheng AM, McGlade CJ. Caspase-dependent cleavage of the hematopoietic specific adaptor protein Gads alters signalling from the T cell receptor. Oncogene. 2001;20:1203–1211. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous