

Profiling of temperature-induced changes in Borrelia burgdorferi gene expression by using whole genome arrays
- PMID: 12654782
- PMCID: PMC152086
- DOI: 10.1128/IAI.71.4.1689-1705.2003
Profiling of temperature-induced changes in Borrelia burgdorferi gene expression by using whole genome arrays
Abstract
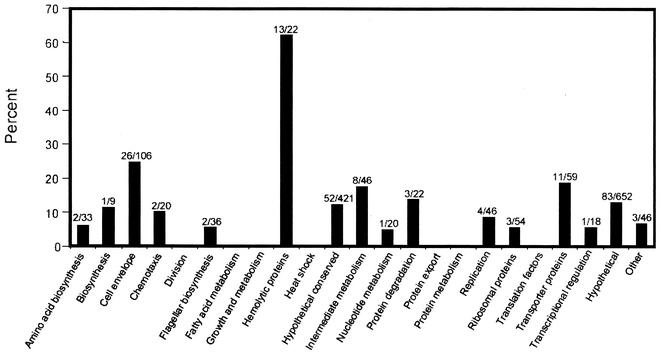
Borrelia burgdorferi is the etiologic agent of Lyme disease, the most prevalent arthropod-borne disease in the United States. The genome of the type strain, B31, consists of a 910,725-bp linear chromosome and 21 linear and circular plasmids comprising 610,694 bp. During its life cycle, the spirochete exists in distinctly different environments, cycling between a tick vector and a mammalian host. Temperature is one environmental factor known to affect B. burgdorferi gene expression. To identify temperature-responsive genes, genome arrays containing 1,662 putative B. burgdorferi open reading frames (ORFs) were prepared on nylon membranes and employed to assess gene expression in B. burgdorferi B31 grown at 23 and 35 degrees C. Differences in expression of more than 3.5 orders of magnitude could be readily discerned and quantitated. At least minimal expression from 91% of the arrayed ORFs could be detected. A total of 215 ORFs were differentially expressed at the two temperatures; 133 were expressed at significantly greater levels at 35 degrees C, and 82 were more significantly expressed at 23 degrees C. Of these 215 ORFs, 134 are characterized as genes of unknown function. One hundred thirty-six (63%) of the differentially expressed genes are plasmid encoded. Of particular interest is plasmid lp54 which contains 76 annotated putative genes; 31 of these exhibit temperature-regulated expression. These findings underscore the important role plasmid-encoded genes may play in adjustment of B. burgdorferi to growth under diverse environmental conditions.
Figures




References
-
- Akins, D. R., S. F. Porcella, T. G. Popova, D. Shevchenko, S. I. Baker, M. Li, M. V. Norgard, and J. D. Radolf. 1995. Evidence for in vivo but not in vitro expression of a Borrelia burgdorferi outer surface protein F (OspF) homologue. Mol. Microbiol. 18:507-520. - PubMed
-
- Benjamini, Y., and Y. Hochberg. 1995. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 57:289-300.
Publication types
MeSH terms
Substances
Grants and funding
- AI 07364/AI/NIAID NIH HHS/United States
- R01 AI027044/AI/NIAID NIH HHS/United States
- T32 AI007364/AI/NIAID NIH HHS/United States
- R01 AI042345/AI/NIAID NIH HHS/United States
- R01 AI037248/AI/NIAID NIH HHS/United States
- AI 45801/AI/NIAID NIH HHS/United States
- AI 37248/AI/NIAID NIH HHS/United States
- P20 RR015564/RR/NCRR NIH HHS/United States
- R56 AI027044/AI/NIAID NIH HHS/United States
- R37 AI027044/AI/NIAID NIH HHS/United States
- AI 27044/AI/NIAID NIH HHS/United States
- R01 AI045801/AI/NIAID NIH HHS/United States
- AI 29735/AI/NIAID NIH HHS/United States
- R56 AI029735/AI/NIAID NIH HHS/United States
- RR 15564/RR/NCRR NIH HHS/United States
- R01 AI029735/AI/NIAID NIH HHS/United States
- AI 42345/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
