

Preorganization of molecular binding sites in designed diiron proteins
- PMID: 12655072
- PMCID: PMC152997
- DOI: 10.1073/pnas.0730771100
Preorganization of molecular binding sites in designed diiron proteins
Abstract
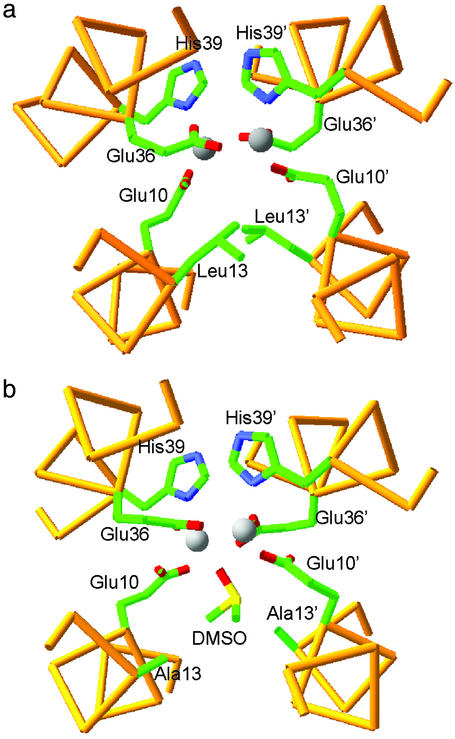
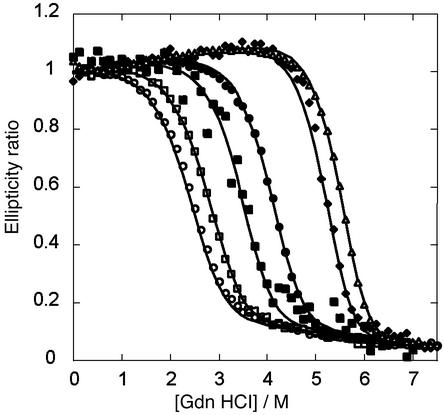
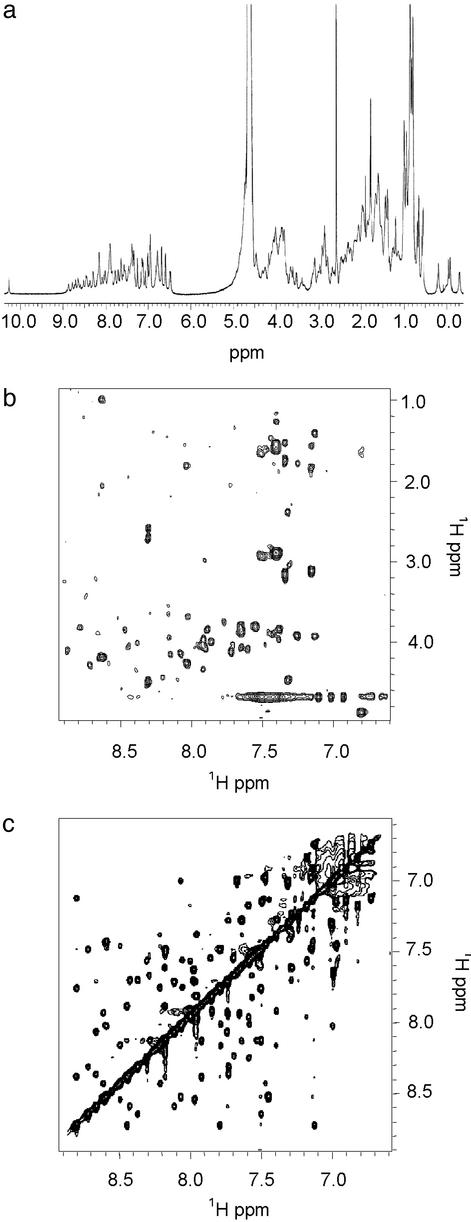
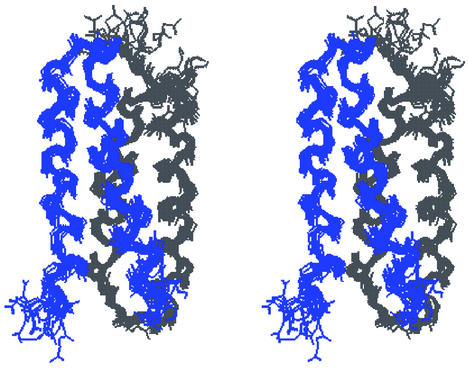
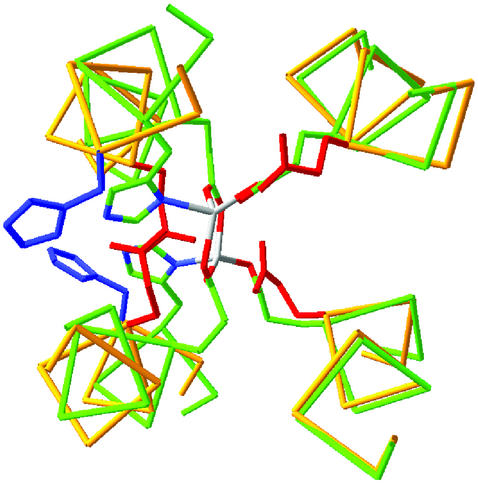
De novo protein design provides an attractive approach to critically test the features that are required for metalloprotein structure and function. Previously we designed and crystallographically characterized an idealized dimeric model for the four-helix bundle class of diiron and dimanganese proteins [Dueferri 1 (DF1)]. Although the protein bound metal ions in the expected manner, access to its active site was blocked by large bulky hydrophobic residues. Subsequently, a substrate-access channel was introduced proximal to the metal-binding center, resulting in a protein with properties more closely resembling those of natural enzymes. Here we delineate the energetic and structural consequences associated with the introduction of these binding sites. To determine the extent to which the binding site was preorganized in the absence of metal ions, the apo structure of DF1 in solution was solved by NMR and compared with the crystal structure of the di-Zn(II) derivative. The overall fold of the apo protein was highly similar to that of the di-Zn(II) derivative, although there was a rotation of one of the helices. We also examined the thermodynamic consequences associated with building a small molecule-binding site within the protein. The protein exists in an equilibrium between folded dimers and unfolded monomers. DF1 is a highly stable protein (K(diss) = 0.001 fM), but the dissociation constant increases to 0.6 nM (deltadeltaG = 5.4 kcalmol monomer) as the active-site cavity is increased to accommodate small molecules.
Figures
References
-
- Honig B. & Yang, A.-S. (1995) Adv. Protein Chem. 46, 27-58. - PubMed
-
- Baldwin E. P. & Matthews, B. W. (1994) Curr. Opin. Biotechnol. 5, 396-402. - PubMed
-
- Lippard S. J. & Berg, J. M., (1994) Principles of Bioinorganic Chemistry (Univ. Sci. Books, Mill Valley, CA).
-
- Holm R. H., Kennepohl, P. & Solomon, E. I. (1996) Chem. Rev. (Washington, D.C.) 96, 2239-2314. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
