

Coexistence of excitatory and inhibitory GABA synapses in the cerebellar interneuron network
- PMID: 12657660
- PMCID: PMC6742031
- DOI: 10.1523/JNEUROSCI.23-06-02019.2003
Coexistence of excitatory and inhibitory GABA synapses in the cerebellar interneuron network
Abstract
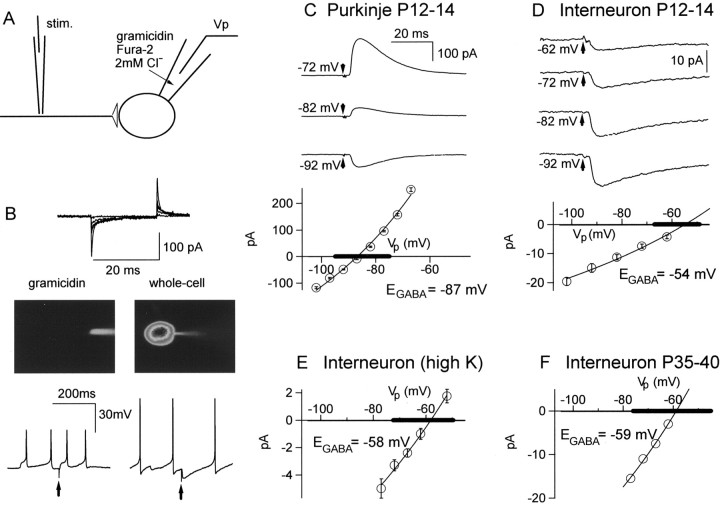
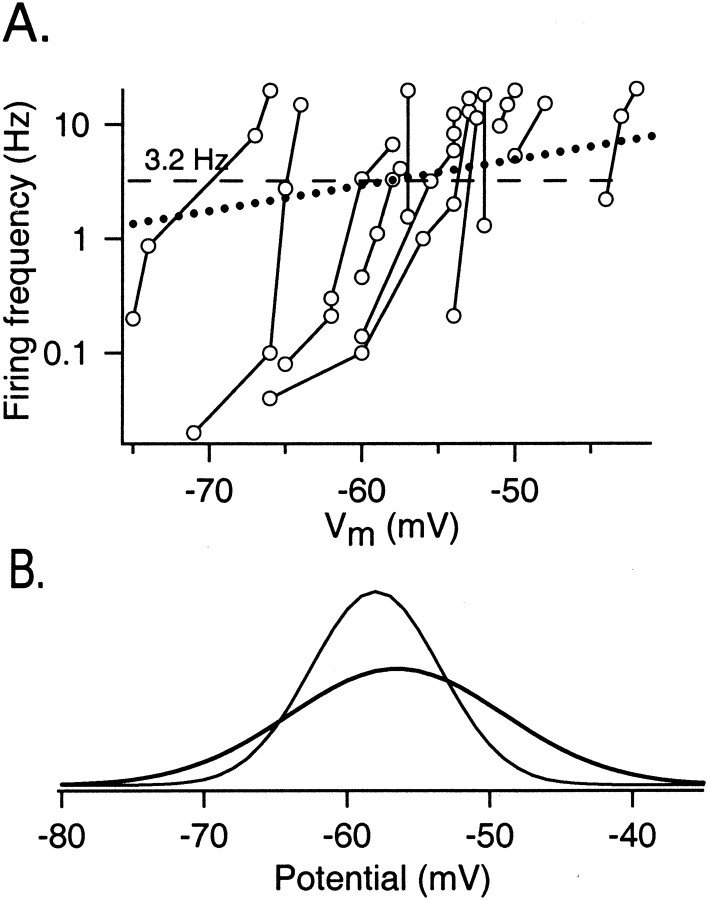
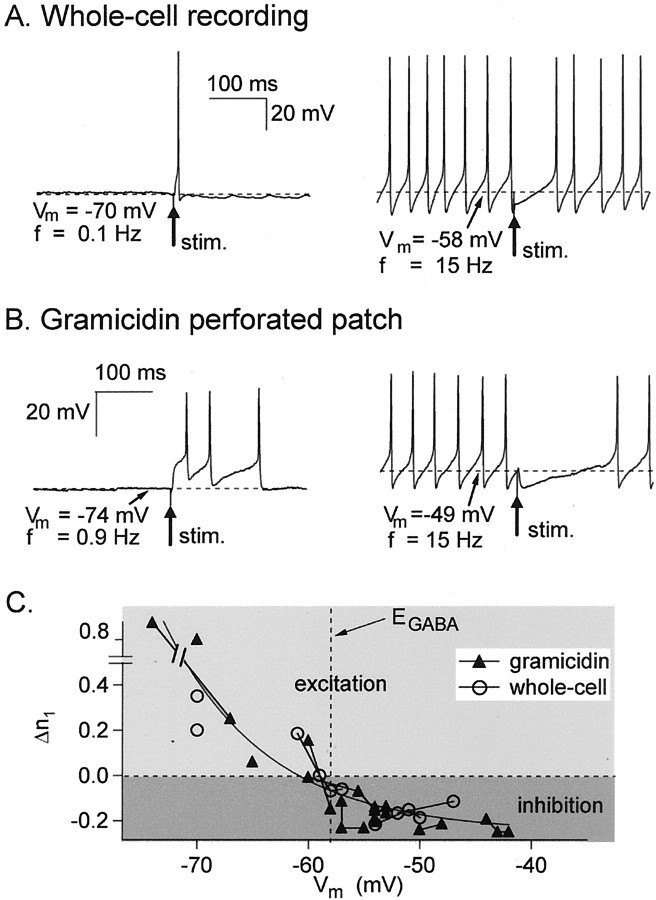
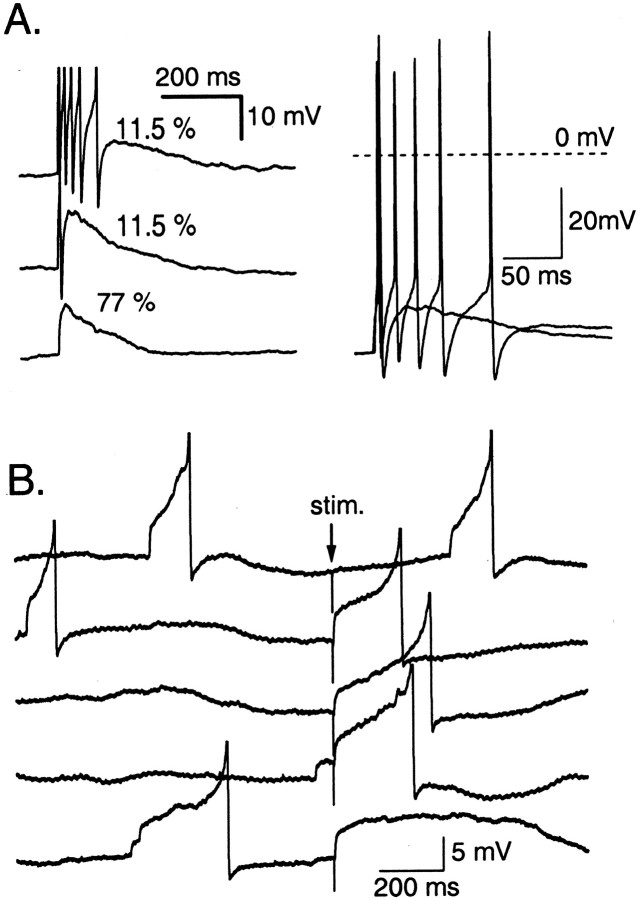
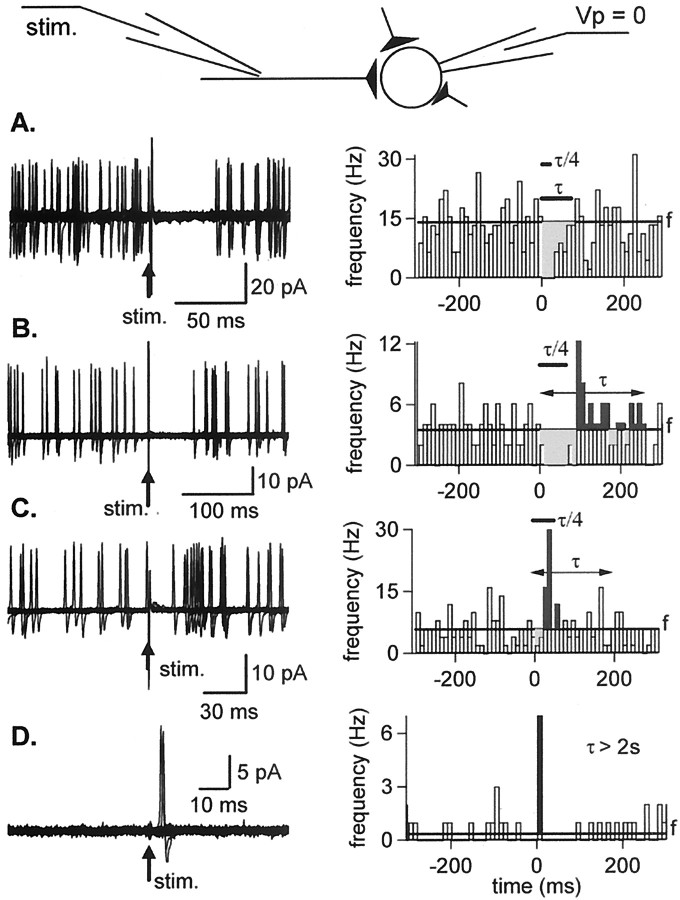
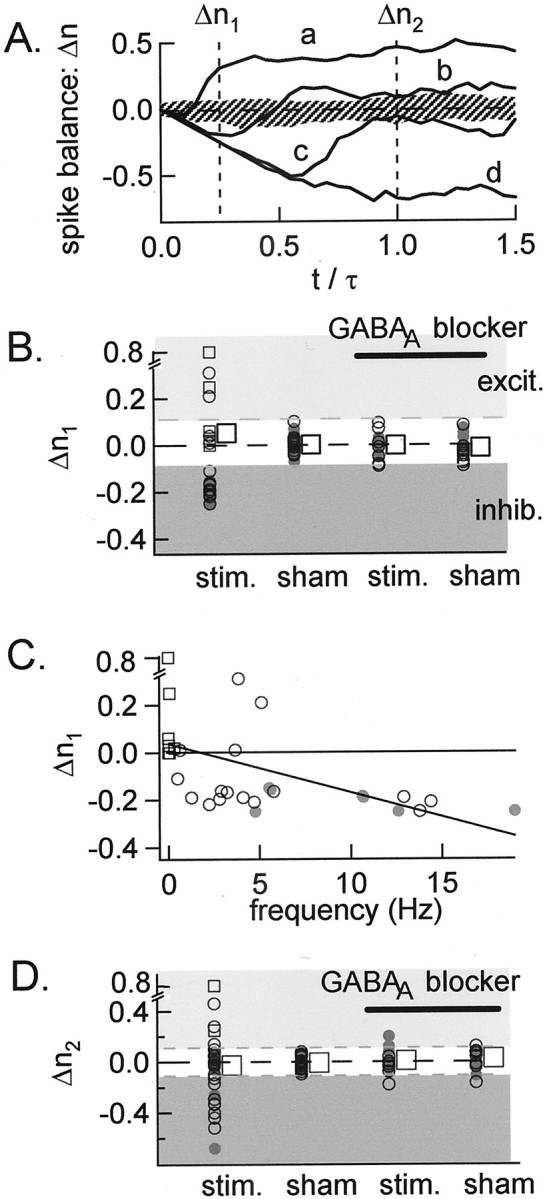
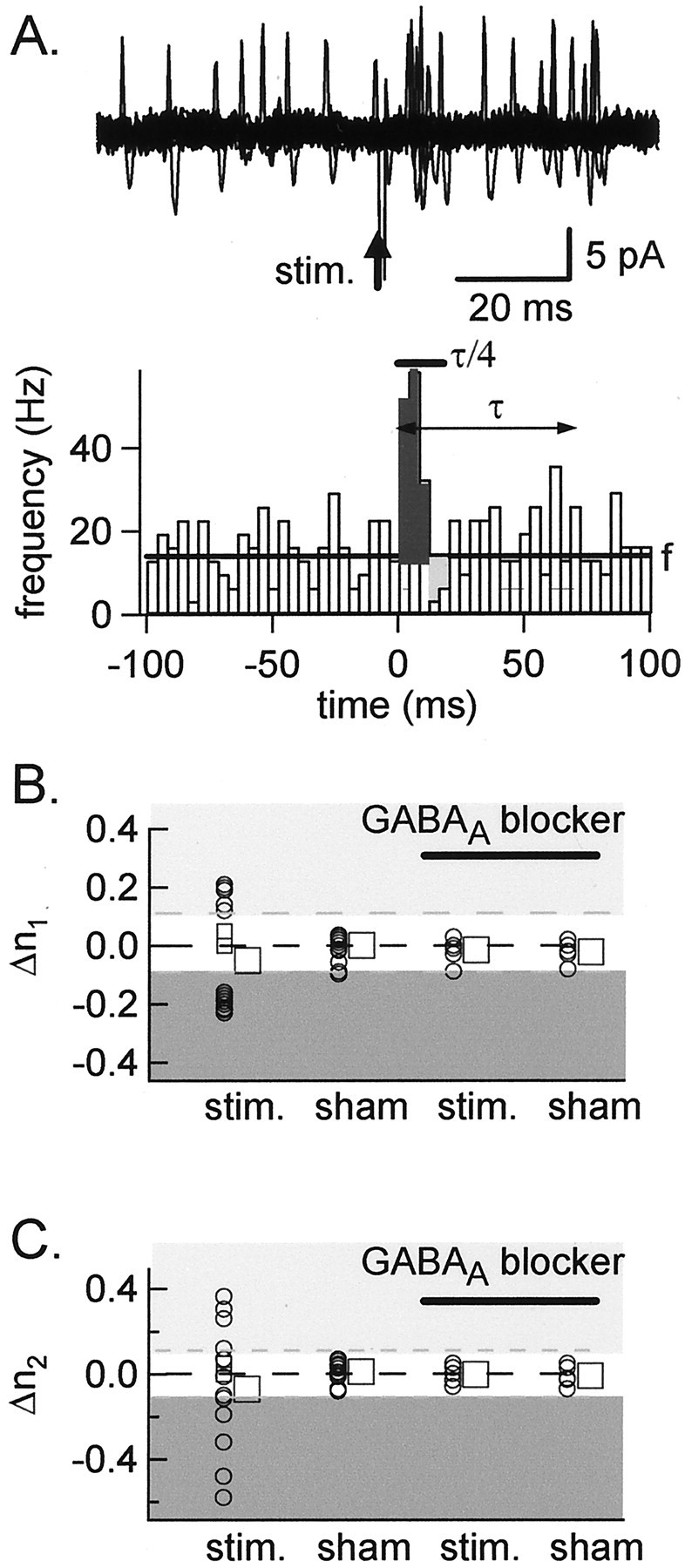
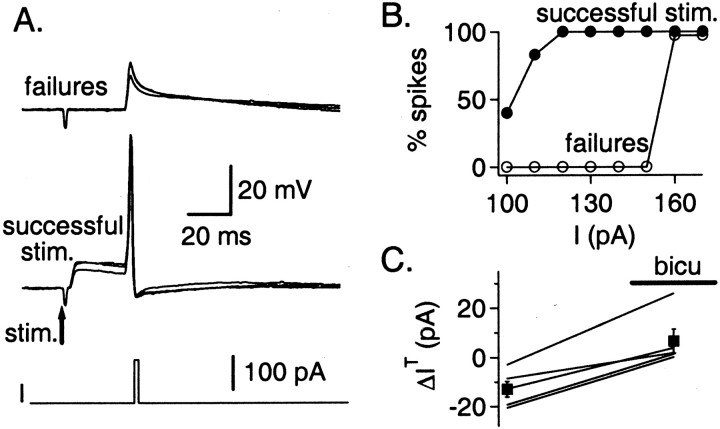
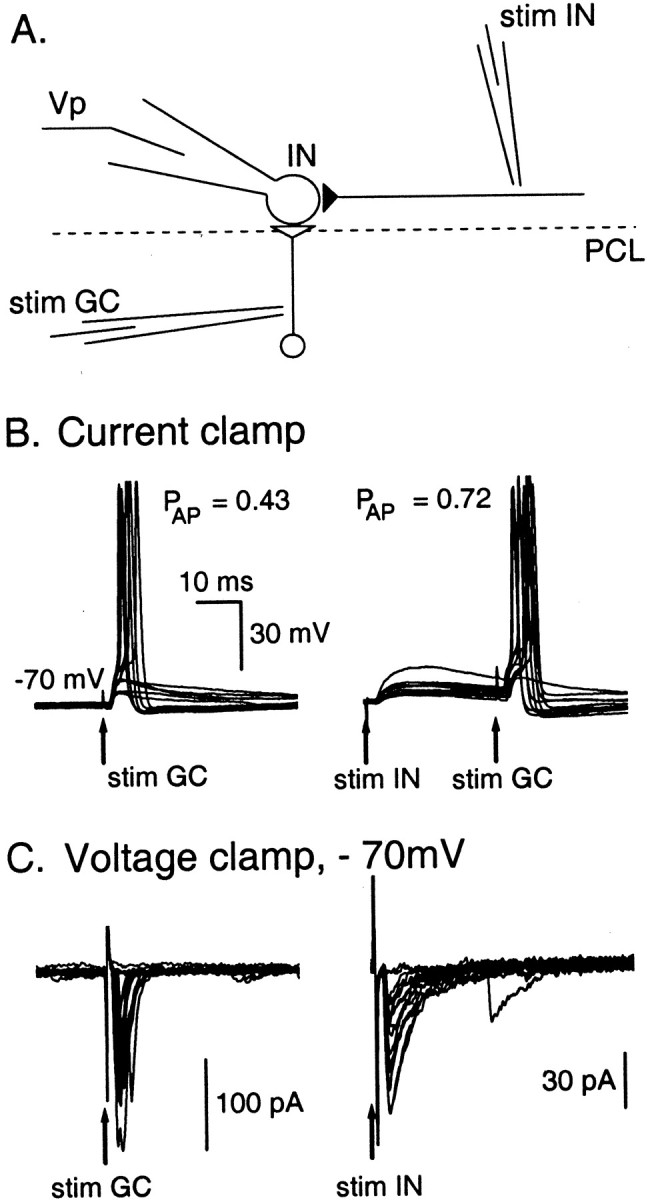
Functional GABA synapses are usually assumed to be inhibitory. However, we show here that inhibitory and excitatory GABA connections coexist in the cerebellar interneuron network. The reversal potential of GABAergic currents (E(GABA)) measured in interneurons is relatively depolarized and contrasts with the hyperpolarized value found in Purkinje cells (-58 and -85 mV respectively). This finding is not correlated to a specific developmental stage and is maintained in the adult animal. E(GABA) in interneurons is close to the mean membrane potential (-56.5 mV, as measured with a novel "equal firing potential" method), and both parameters vary enough among cells so that the driving force for GABA currents can be either inward or outward. Indeed, using noninvasive cell-attached recordings, we demonstrate inhibitory, excitatory, and sequential inhibitory and excitatory responses to interneuron stimulation [results obtained both in juvenile (postnatal days 12-14) and subadult (postnatal days 20-25) animals]. In hyperpolarized cells, single synaptic GABA currents can trigger spikes or trains of spikes, and subthreshold stimulations enhance the responsiveness to subsequent excitatory stimulation over at least 30 msec. We suggest that the coexistence of excitatory and inhibitory GABA synapses could either buffer the mean firing rate of the interneuron network or introduce different types of correlation between neighboring interneurons, or both.
Figures
References
-
- Aizenman CD, Linden DJ. Regulation of the rebound depolarization and spontaneous firing patterns of deep nuclear neurons in slices of rat cerebellum. J Neurophysiol. 1999;82:1697–1709. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources